Chapitre 7 : Une impossible synthèse
Ce que nous disent les programmes officiels d’enseignement en SVT
« La théorie de l’évolution est elle-même encore
très jeune et il est par conséquent hors de doute que les recherches
ultérieures doivent modifier très notablement les idées actuelles, y compris
celles qui sont strictement darwiniennes, sur la marche de l’évolution des
espèces »
Engels. Anti-Dühring
La biologie officielle lutte sur deux fronts. Sur l’un,
elle doit faire face à une crise théorique qui l’assiège avec la dernière
vigueur (chapitres 2 et 3). Sur l’autre, il lui faut sans cesse réaffirmer ses
positions contre l’imminence d’une synthèse matérialiste de plus en plus facile
à effectuer (chapitres 5 et 6) … Synthèse dont elle fournit bien malgré elle
toutes les clefs !
Manifestement le néodarwinisme n’a plus l’initiative
dans cette « guerre »… Comment peut-il tenir sur ces deux fronts sans
perdre la face ? Comme nous l’avons déjà montré, il a perdu d’avance, et
nous n’irons pas chercher bien loin la preuve de cet échec annoncé ; elle
saute aux yeux des enseignants du second degré qui ont suivis les changements
de programme depuis les années quatre-vingt. A chaque réforme, le retard de
l’enseignement sur la recherche s’amenuise : Dans tous les domaines de la
biologie les contradictions s’intensifient et la recherche en expose
instantanément toutes les impasses, pendant que l’enseignement piétine, pris en
tenaille entre la nécessité d’une métamorphose et l’impossibilité
d’officialiser des théories alternatives trop dissidentes… Il est hors de
question d’enseigner aux élèves autre chose qu’une science sûre d’elle même,
assise sur des bases solides et objectives ! Notons par exemple que les
manuels de SVT se distinguent fondamentalement des manuels de Physique-Chimie
par l’absence absolue de références à l’histoire des sciences ! (Ceux de
Physique-Chimie au contraire en font un usage massif et régulier).
Dans ce contexte, rédiger les nouveaux programmes de
Sciences de
La démonstration s’appuiera essentiellement sur une
comparaison des anciens et nouveaux programmes de la filière scientifique en
Sciences de
A.
Réécrire l’histoire
a.
Ignorer Darwin…
L’histoire des sciences n’a jamais été enseignée en SVT
au lycée, et pour cause… Il n’est pas question de gaspiller le précieux temps
des futurs bacheliers avec la question des idéologies scientifiques ! D’un
manuel scolaire à l’autre, on trouvait parfois dans les années quatre-vingt,
des hagiographies sur Mendel ou Morgan, rarement sur Darwin. Mais les programmes
officiels d’alors n’y faisaient jamais référence[1].
Il y a plus étonnant concernant Darwin. Si le
mendélo-morganisme est enseigné depuis les années soixante dix, la science de
l’évolution, question centrale de la biologie depuis la fin du XIXe
siècle, n’apparaît dans les programmes que dans les années quatre-vingt dix.
Elle n’est abordée pratiquement que depuis 2001… Aujourd’hui paradoxalement,
les anecdotes concernant les grands découvreurs ont totalement déserté les
manuels scolaires. Plus les notions scientifiques sont affirmées, plus leurs
auteurs sont effacés[2].
C’est hors du lycée que les élèves ou les étudiants
découvriront comment Darwin a révolutionné la biologie ! Sa théorie elle
même ne s’est pas implantée dans les programmes sans difficultés. Totalement
ignorée dans les textes antérieurs, elle apparaît timidement dans les années
quatre-vingt dix.
|
Ancien programme de TS (B.O. hors série n° 6 du 9 juin 1994) |
Nouveau programme de TS (B.O. hors série n° 5 du 30 août 2001) |
|
En
initiant aux mécanismes, encore
largement hypothétiques, de l’évolution biologique, [cette partie de
programme] aide à comprendre la dynamique de la biodiversité. (…) L’évolution est la seule explication
scientifique qui rende compte des constats : unité, diversité du
monde vivant et changements ayant lieu au cours des temps géologiques. L’évolution
implique une filiation entre les espèces ; les espèces actuelles
dérivent d’ancêtres communs plus ou moins éloignés dans le temps ; une
origine commune à toutes les espèces est hautement probable. |
Les êtres
vivants partagent des propriétés communes (structure cellulaire, ADN,
modalités de la réplication et de l’expression des gènes, code génétique).
Ces propriétés traduisent
une origine commune. L’état
actuel du monde vivant résulte
de l’évolution. Toutes les espèces vivantes actuelles et toutes les
espèces fossiles sont
apparentées mais elles le sont plus ou moins étroitement. |
Avant d’être un fait scientifique, l’évolution n’était
donc qu’une « hypothèse hautement probable »… En revanche la manière
d’amener cette notion a quelque peu changé. Dans l’ancien programme on insiste
sur l’unité du monde vivant aux niveaux morphologique, embryologique et
moléculaire, impliquant l’origine
commune des espèces actuelles. Dans le nouveau programme, c’est la diversité
des espèces actuelles et anciennes qui prévaut, impliquant des relations de
parenté hiérarchisées entre elles. Le cladisme, suggéré dans l’ancien programme
par une phrase finalement effacée lors des allègements à la fin des années quatre-vingt
dix, est aujourd’hui la méthode reine de mise en évidence de
l’évolution.
Le cladisme est une méthode formaliste assez récente
qui quantifie les caractères homologues « dérivés » (comprendre
« évolués ») partagés ou non au sein d’un ensemble donné d’espèces
actuelles. Le « principe de parcimonie » commande le choix de l’arbre
phylogénétique le plus probable parmi toutes les hiérarchies de parenté
possibles entre les espèces de ce groupe ; on retient l’arbre qui suppose
le plus petit nombre d’innovations successives, l’apparition simultanée d’une
même innovation structuralo-fonctionnelle sur des branches évolutives
indépendantes étant considérée comme fortement improbable[3].
Par conséquent, les nœuds réunissant plusieurs branches
ne correspondent plus à des « ancêtres communs » (ancien programme)
mais à des « ancêtres communs théoriques ». De plus, l’imperfection
des restes fossiles connus, les inévitables lacunes correspondant aux espèces
fossiles non encore découvertes, le caractère subjectif enfin, du choix des
caractères homologues utilisés, impose à cette méthode un relativisme assumé.
|
Ancien programme de TS |
Nouveau programme de TS |
|
Des
relations de parenté entre les êtres vivants peuvent être proposées à partir
de l’étude de caractéristiques morphologiques, anatomiques et embryologiques
d’organismes actuels et fossiles. La
recherche des parentés s’appuie également sur des comparaisons, au niveau
moléculaire, de séquences de gènes homologues ou de produits de l’expression
de ces gènes. |
L’établissement
de relations de parenté entre les vertébrés actuels s’effectue par
comparaison de caractères homologues (embryonnaires, morphologiques,
anatomiques et moléculaires). Les comparaisons macroscopiques prennent en
compte l’état ancestral et l’état dérivé des caractères. Seul le
partage d’états dérivés des caractères témoigne d’une étroite parenté. Ces
relations de parenté contribuent à construire des arbres phylogénétiques. Les
ancêtres communs représentés sur les arbres phylogénétiques sont hypothétiques, définis
par l’ensemble des caractères dérivés partagés par des espèces qui leur sont
postérieures ; ils ne correspondent pas à des espèces fossiles précises.
Une espèce fossile ne
peut être considérée comme la forme ancestrale à partir de laquelle se
sont différenciées les espèces postérieures. |
Du point de vue philosophique, nous savons à quel point
il est facile de passer d’un relativisme aussi fécond qu’inévitable, à un
subjectivisme particulièrement délétère en science. Ainsi, l’enseignant affirme
que l’évolution est un fait, mais glisse très facilement du caractère
éternellement hypothétique de toute construction cladiste au caractère fondamentalement hypothétique de la notion
même d’évolution : certes c’est un fait scientifique, mais il est indémontrable…
Ernst Mayr lui même ne s’y est pas trompé. Pour lui
comme pour la majorité de ses collègues néodarwiniens, le cladisme, produit
d’importation de l’« idéalisme allemand » destiné à contrarier
l’« empirisme anglo-saxon », doit être rejeté. Cette polémique est
d’ailleurs loin d’être close ; le cladisme doit être rejeté parce qu’il
donne de l’évolution une image subjectiviste, abstraite et finalement
anti-darwinienne, au sens où il passe sous silence la notion primordiale
d’adaptation des espèces à leur milieu. Cette omission n’est pas fortuite. Dans
un premier temps, les auteurs du programme se réclament ouvertement de Darwin
lorsqu’ils avancent la théorie cladiste, même si ce dernier n’est pas cité dans
le programme. Une conférence préparatoire[4]
au sujet du nouveau programme en
fait état : « La
cladistique est un « enfant de Darwin ». On lit dans l’Origine des
Espèces « Le système naturel est fondé sur le principe de
descendance avec modification. Les caractères que les naturalistes considèrent
comme montrant une pré-affinité entre deux ou plusieurs espèces sont ceux qui
ont été hérités d’un parent commun par conséquent toute la
classification est généalogique. La communauté de descendance est le lien caché
que les naturalistes ont cherché inconsciemment et non quelques plans inconnus
de création. » Dans cette citation de Darwin, on retrouve les notions de
parenté, de temps, de mort des espèces et d’ancêtre hypothétique commun. Le
problème est de passer du concept de généalogie au concept de parenté (pas de
confusion entre généalogie et lien de parenté). » Autrement dit, on
cite un texte anti-finaliste de Darwin qui n’a rien à voir avec
l’indéterminisme cladiste, on en tire la notion clef de généalogie, puis on
déclare que le cladisme équivaut au rejet même de cette notion !
Dans un second temps, et parallèlement à
l’officialisation du cladisme, méthode formaliste et indéterministe, les
notions polémiques de sélection naturelle, de compétition et de spéciation
allopatrique disparaissent[5].
|
Ancien programme de TS |
Nouveau programme de TS |
|
Les
innovations génétiques peuvent se traduire ou non dans le phénotype. Si
certaines mutations sont neutres,
d’autres s’expriment et peuvent même avoir des conséquences importantes,
surtout si ce sont des gènes
du développement qui sont touchés. La sélection naturelle,
s’exerçant sur des populations soumises à des conditions de milieu
différentes, privilégie la conservation des allèles ou associations
alléliques favorables dans les conditions écologiques du moment. (exploitation
de documents relatifs à un exemple ancien (concurrence édentés-autres mammifères lors de la
liaison Amérique du nord – Amérique du sud, concurrence marsupiaux – placentaires au
quaternaire). (…) La spéciation ou naissance
d’espèces nouvelles à partir d’une espèce mère implique l’isolement reproductif
entre les populations de l’espèce mère considérée. |
Les
innovations génétiques sont aléatoires et leur nature ne dépend pas des caractéristiques du
milieu. (…) Les
innovations génétiques peuvent être favorables, défavorables ou neutres pour la survie
de l’espèce. Parmi les
innovations génétiques seules celles qui affectent les cellules germinales d’un
individu peuvent avoir un impact évolutif. Les
mutations qui confèrent un avantage sélectif aux individus qui en sont
porteurs ont une probabilité plus grande de se répandre dans la population. Des
mutations génétiques peuvent se répandre dans la population sans conférer
d’avantage sélectif particulier (mutations dites neutres). Des
mutations affectant les gènes
du développement peuvent avoir des répercussions sur la chronologie et
la durée relative de la mise en place des caractères morphologiques. De
telles mutations peuvent avoir des conséquences importantes. |
Si on retrouve la notion d’avantage sélectif, celle de
sélection naturelle n’est plus identifiable. La sélection naturelle s’exerçait
sur des populations comme action conservatrice (ectogenèse), ce sont
aujourd’hui les mutations qui agissent sur des individus en leur conférant un
« avantage sélectif » (autogenèse) ! Dominés par le neutralisme,
les mécanismes darwiniens de l’évolution laissent une large place à la
variabilité individuelle aléatoire et finalement à l’autogenèse indéterministe,
tout en réaffirmant les notions weismanniennes de lignée germinale et
d’indépendance vis à vis du milieu.
La recherche aurait-elle été trop rapide ? A peine
l’évolution biologique fait-elle irruption dans les programmes que Darwin est
déjà renvoyé au placard ! Amputée de sa notion de sélection naturelle, la
théorie darwinienne subsiste dans les termes d’une variabilité individuelle qui
intéresse la génétique plus que la science de l’évolution. On censure donc le
problème de l’adaptation des organismes à leur milieu : Désormais divorcée
de la génétique des populations, la génétique moléculaire assumera seule la
tâche d’expliquer l’évolution biologique. C’est à force de taire son inaptitude
à remplir cette mission que les notions hétérodoxes des post-néodarwiniens
avanceront masquées dans les programmes.
b.
… Honorer Mendel et Morgan
Le programme de spécialité ne consacre pas moins de dix
semaines, soit un tiers de l’année scolaire, au chapitre d’histoire des
sciences intitulé « Des débuts de la génétique aux enjeux actuels des
bio-technologies ». Pourtant, à la différence des deux autres thèmes de
spécialité, celui ci ne comporte dans le Bulletin Officiel aucune introduction,
aucun texte explicatif préalable. Mieux ; aucun lien avec un quelconque
chapitre du programme de tronc commun, aucune transversalité avec les programmes
d’histoire ou de philosophie enseignés[6]
en Terminale S.
|
Thème 2. Des débuts de la génétique aux enjeux actuels
des biotechnologies (10 semaines) |
|
|
Activités envisageables |
Notions et contenus |
|
Réalisation
d’une dissection florale en relation avec la technique expérimentale de
Mendel. Observation d’un fruit et d’une graine. Analyse
d’expériences relatives au monohybridisme et au dihybridisme dans la
perspective des travaux de Mendel. Constat du
parallélisme entre le comportement des chromosomes et celui des facteurs
héréditaires. Etude des
résultats de croisement chez la drosophile dans le cas de l’hérédité liée au
sexe et interprétation des résultats dans le cadre de la théorie
chromosomique. Réflexion sur la valeur heuristique d’une
théorie scientifique. Localisation
de trois gènes sur un chromosome à partir de données expérimentales Digestion
de l’ADN par des enzymes de restriction et électrophorèse. (…) |
Les début
de la génétique : Les travaux de Mendel (1870). Les
travaux de Mendel reposent sur une analyse quantitative d’expériences d’hybridation chez les
plantes. Novateurs dans leur méthodologie, ces travaux visaient à obtenir des
hybrides stables. Dans une contexte scientifique où les gènes n’étaient pas
connus, ils ont apporté une rupture conceptuelle : a.
réfutation de la notion d’hérédité par mélange, b.
introduction du concept d’hérédité particulaire avec ségrégation
indépendante des facteurs héréditaires. La
compréhension des travaux de Mendel repose sur la connaissance des principes
de la reproduction sexuée des végétaux. La théorie
chromosomique de l’hérédité. La
redécouverte des lois de Mendel et les découvertes dans le domaine de la
cytologie à la fin du XIXe siècle conduisent à l’émission de la
théorie chromosomique de l’hérédité (1903) par deux cytologistes et à l’invention du mot gène. Les
travaux de Morgan sur L’avènement
de la biologie moléculaire : Une nouvelle rupture. La nature
chimique du gène (ADN-double hélice), la relation gène-protéine, les modalités de
l’expression génétique, notions déjà étudiées dans les programmes de seconde
et de première, doivent être replacées dans une perspective historique. Elles ne sont pas au
programme en tant que telles. La
révolution technologique du début des années 70. L’utilisation
des enzymes de restriction ouvre la voie du clonage des gènes et de leur
séquençage. En contribuant à une évolution importante du concept de gène et de la
perception du polymorphisme, elle fait entrer la génétique dans l’ère des
biotechnologies. (…) [OGM,
dépistage, thérapie génique] |
Incongruité flagrante, simplisme suspect, voilà un
chapitre particulièrement étrange aux yeux des élèves comme à ceux des
enseignants. En somme, Mendel et Morgan n’auront jamais été si honorés dans les
lycées que depuis le désaveu public du mendélo-morganisme dans la presse
scientifique et les laboratoires de recherche !
Remarquons ici deux points importants. 1) Le choix des
hommes à qui on rend hommage, à travers un récit totalement néo-positiviste. 2)
La réactualisation de quelques notions fondamentales de la génétique
moléculaire, réinvesties, rebaptisées, révisées même selon le goût du jour.
Constatons tout d’abord qu’aucun généticien critique
voire dissident vis à vis du mendélo-morganisme n’est cité dans ce récit. Par
suite, pas un des manuels scolaires publiés depuis n’y fera référence[7].
Aucune polémique donc, exceptée celle que lança Mendel lui même contre les
partisans de l’« hérédité par mélange ». Rien sur Goldschmidt, rien
sur Mac Clintock… rien sur le problème des macromutations, pourtant omniprésent
depuis le programme de seconde jusqu’à celui de TS !
Si les travaux de Mendel marquent une
« rupture » dans l’histoire des conceptions de l’hérédité, Morgan en
sera le continuateur triomphant. En revanche, on ne comprend pas bien en
quoi la
génétique
moléculaire marque une
« nouvelle rupture » par rapport au mendélo-morganisme ! Cette
étonnante perspective se rapporte évidemment au schéma du paradigme
kuhnien ; La notion de rupture est aujourd’hui incontournable en histoire
des sciences, mais la biologie souffre d’un grave déphasage par rapport à l’épistémologie
moderne… Les auteurs de ce programme tiennent à annoncer ces
« ruptures » comme garantie de scientificité, mais ils ne peuvent
pour autant se résoudre à relativiser leur mendélo-morganisme à l’heure où il
trône encore au sommet de l’édifice scientifique officiel.
C’est une contradiction implicite qui amène les auteurs
à offrir les gages d’une humilité scientifique propre à séduire les
sensibilités indéterministes alors même qu’ils tentent de réaffirmer
brutalement, maladroitement même, les piliers conceptuels du
mendélo-morganisme : formalisme (« analyse quantitative ») et
réductionnisme idéaliste (« notion de gène »)… La « perspective
historique » à travers laquelle on valorise les travaux successifs de
Mendel, Morgan, Monod est en réalité celle d’un progrès continu ouvrant la voie
aux « triomphes » récents de la biotechnologie… Si cette position
tout à fait positiviste s’accommode mal du processus cyclique paradigme /
révolution scientifique cher à Thomas Kuhn, c’est la « valeur
heuristique » des théories scientifiques qui sauve sinon leur objectivité,
du moins leur authenticité scientifique ; une théorie est condamnée à
rester subjective mais elle a toujours une valeur explicative qui justifie sa
légitimité. Tel est le néo-positivisme maladroit du mendélo-morganisme proposé
aux élèves.
Par quelques détours sémantiques, le déterminisme
réductionniste « un gène-un trait » devient une très humble
« hérédité particulaire », formule minimale mais finalement toujours
réductionniste, le dogme central devient une timide « relation
gène-protéine », dont on tait l’unilatéralité polémique, la notion de gène
enfin n’est plus brandie comme une explication totale, mais devient de façon
restrictive et empirique, une « unité de fonction, de recombinaison, de mutation »…
impossible à taire, l’unité de fonction est alors diluée, mais nous ne sommes
pas dupes. Aucune de ces trois redéfinitions empiriques ne tient plus
aujourd’hui (cf. chapitre 1)…
Peut-on concéder finalement cet échec théorique ?
Oui, à condition qu’un tel échec soit dénué de conséquence : ainsi, la
génétique moléculaire « contribue à une évolution importante du
concept de gène ». Ceci est à la fois une contre-vérité historique, si on
passe Kupiec et Sonigo sous silence, et une flagrante hypocrisie ; aucun
manuel scolaire n’appuie, ne fut-ce que par un seul exemple anecdotique, cette
notion exigible du programme ! Extrême prudence : Les élèves doivent
quitter le lycée avec l’idée que la notion de gène a fortement évolué, mais il
faut absolument leur cacher en quoi consiste cette évolution… La présente
analyse nous évitera d’être étonnés d’une telle inconséquence.
On remarquera ici l’absence des termes jadis
surexploités de programme génétique, de dogme central (le mot
« dogme » passe très mal aujourd’hui !), d’information
génétique, de déterminisme génotypique, … Les auteurs s’en tirent à bon compte.
Le document d’accompagnement au programme de TS, réservé aux enseignants,
commente : « C’est un moment privilégié pour aborder la nature des
théories scientifiques : ce ne sont pas des réalités découvertes, mais des
constructions intellectuelles qui reflètent l’idée que l’on se fait de la
réalité à un moment donné de l’histoire des sciences. Les concepts
scientifiques sont réfutables ; ils sont confrontés en permanence aux
faits d’observation et aux résultats expérimentaux. La science construit son
objet d’étude et ne se contente pas d’accumuler des observations. » La
chose est donc annoncée ; nous nageons dans le poppérisme le plus
vulgaire, oubliant au passage que Popper s’est précisément employé à démonter
la notion d’évolution biologique pour construire sa théorie de la
réfutabilité !
Si on se place du point de vue du lycéen, le récit
proposé offre tous les arguments du mendélisme qui furent lumineusement
confirmés puis élargis par Morgan, par Monod, jusqu’à jeter les bases d’une
technologie efficace au plus haut point, tout en avançant une version humble et
relativiste du progrès scientifique au niveau conceptuel. Ignorant tout des
réelles polémiques qui firent régulièrement et justement obstacle au
mendélo-morganisme, comment notre lycéen peut-il ne pas être séduit par cette
science univoque qui présente son triomphe avec une extrême modestie
qu’apparemment rien ne justifie, qui se donne tant de mal à relativiser des découvertes
dont l’objectivité est si flagrante ! Voilà un habile tour de passe-passe…
qui peut encore tenir le temps que le mendélo-morganisme achève son
agonie !
Si ce chapitre d’histoire des sciences est une manne
pour notre analyse, il ne faut pas oublier pour autant qu’il est marginal. Il
l’est dans le programme de spécialité, dont la plupart des élèves de terminale,
non-spécialistes, ne bénéficient pas. Il l’est plus encore vis à vis du
programme de tronc commun qui précisément se distingue par une scrupuleuse
discrétion idéologique. Avançons néanmoins que ce dernier répond à grande
échelle et implicitement, aux axes qui déterminent le présent chapitre ;
éviter toute contre-argumentation ouverte à la théorie néodarwiniste d’une
part, passer en fraude l’indéterminisme par des sous-entendus permanents en
contradiction flagrante avec les anciens programmes d’autre part.
B.
Un pas en avant : L’élan indéterministe
a.
Sur l’évolution
Dès l’ancien programme, la question de l’évolution
biologique passe par une forme non-polémique de la théorie neutraliste. On
envisageait la notion de mutation neutre comme le pivot d’une démonstration
gradualiste : Les gènes évoluent par accumulation de mutations neutres
jusqu’à l’acquisition éventuelle et accidentelle d’une nouvelle fonction. Une
phrase contenant les notions de duplication génique et de gène homéotique[8]
coexistait avec ce modèle orthodoxe comme si la juxtaposition micro-évolution /
macro-évolution n’avait jamais été problématique.
|
Ancien programme de TS |
Nouveau programme de TS |
|
Les
mutations jouent un rôle fondamental et sont à l’origine des différents
allèles d’un gène ; typiquement spontanées, non orientées, elles
interviennent avec une faible fréquence mais peuvent toucher de nombreux
gènes et devenir plus
nombreuses sous l’influence de certains facteurs du milieu. Des duplications géniques peuvent intervenir et
une évolution divergente des duplicata produits peut expliquer l’apparition de gènes nouveaux.
(…) Si
certaines mutations sont neutres,
d’autres s’expriment et peuvent même avoir des conséquences importantes,
surtout si ce sont des gènes
homéotiques qui sont touchés. |
Au sein du
génome d’une espèce, les similitudes entre gènes (familles de gènes) sont interprétées comme
le résultat d’une ou plusieurs duplications d’un gène ancestral. La divergence des gènes
d’une même famille s’expliquent par l’accumulation de mutations. Dans
certains cas, ces processus peuvent conduire à l’acquisition de nouvelles fonctions. Les innovations génétiques sont aléatoires et
leur nature ne dépend pas des caractéristiques du milieu. Des
mutations génétiques peuvent se répandre dans la population sans conférer
d’avantage sélectif particulier (mutations dites neutres). Des
mutations affectant les gènes
homéotiques peuvent
avoir des répercussions sur la chronologie et la durée relative de la mise en place des
caractères morphologiques. |
Le nouveau programme reprend cette juxtaposition, mais
en mettant l’accent sur l’indépendance des innovations génétiques vis à vis du
milieu ; avancée significative de l’indéterminisme donc, malgré la
similitude des notions abordées. Parallèlement, l’alibi neutraliste ouvre
désormais la voie à une officialisation du saltationnisme modéré de Jean
Chaline. Il y est déjà question des (trop) difficiles notions d’hétérochronie
et de macromutation[9]. L’enseignement de la
théorie des « horloges du vivant » que notre renégat saltationniste vient à peine de
réactualiser apparaît comme un pari sur ce que deviendra la théorie de
l’évolution dans un avenir proche. Rappelons que Chaline est le premier à
affirmer que la théorie synthétique n’est pas vraiment néodarwiniste ! La
stratégie des auteurs se précise…
D’une façon générale, l’accent est mis sur le caractère
anti-déterministe des thèses anti-finalistes qu’on suggère aux élèves. Ainsi
par exemple, ce chapitre est l’occasion d’une mise au point digne du plus grand
intérêt. Hervé Le Guyader indique au cours de la conférence citée plus
haut : « Le concept d’échelle des êtres, lié à la notion de
progrès, est toujours présent : il faut démontrer que ce concept est faux.
Exemples d’idées fausses : Les mammifères sont mieux que les reptiles /
Parmi les mammifères, les primates sont les meilleurs / Parmi les primates,
l’Homme est à placer au sommet / … Ce concept d’échelle des êtres entraîne deux
choses : La finalité (tout est fait dans un but donné) contre la notion de
contingence. » Commentant le chapitre concernant la lignée humaine, le
même conférencier tombe plus loin dans un humanisme bioéthique des plus
naïfs : « L’échelle
des êtres est une
source scientifique de racisme. Cela est du à nouveau, à une
confusion entre généalogie et parenté. (…) On ne doit pas
considérer une industrie moins bien qu’une autre [!]» Nous sommes ici dans
le « tout se vaut » condamné faute de dialectique, à faire de
l’antiracisme ethniste et « différencialiste » (conseillant par
exemple très « respectueusement » aux pays économiquement
sous-développés de rester ce qu’ils sont !)… De même, on indique dans le document
d’accompagnement au programme de TS : « Il convient d’aller à
l’encontre d’un certain nombre d’idées fausses (vision linéaire de l’évolution,
finalisme et anthropocentrisme conduisant à décrire l’évolution biologique
comme une série de perfectionnements aboutissant à l’Homme, persistance du
concept de fossile vivant,…). » On retient donc du saltationnisme non
son contenu anti-gradualiste mais ses innovations idéologiques très en vogue
sur le caractère aléatoire de l’évolution et le rejet de la notion de progrès.
Ce document précise que « les notions de convergence et de
réversion », incompatible avec l’évolution aléatoire, comme « la
présentation et la discussion des différentes théories de l’évolution »
ne doivent pas être abordées… Obscurantiste, cette position n’en est donc pas
moins inconfortable ! On note plus loin « Cette partie du
programme est une ouverture intellectuelle [ !]. Elle a pour but
d’envisager des hypothèses explicatives de l’évolution des espèces. La
complexité des relations possibles entre mécanismes de l’évolution et génétique
est telle qu’il est impossible d’en donner une vision d’ensemble ou des
mécanismes explicatifs. » Cruel aveu aux enseignants, qu’on épargnera
sous cette forme un peu brutale aux élèves eux mêmes !
Point intéressant : On voit poindre au delà de
certaines notions officielles indéterministes, des modèles bien plus
polémiques, de type post-néodarwinien… Parmi les sujets proposés aux élèves de
terminale S pour l’évaluation des capacités expérimentales de fin d’année, on
trouve par exemple un thème traitant explicitement des thèses d’Anne
D’ambricourt Malassez. Cette bête noire du néodarwinisme actuel,
antidarwinienne très critique vis à vis de la génétique moléculaire (voir
chapitre 3), est citée dans le contexte des mécanismes de l’hominisation ;
problématique qui se départit désormais des explications réductionnistes
habituelles… En travaux pratiques, les protocoles de comparaison des crânes du
chimpanzé et de l’Homme font référence à la théorie des transformations de
D’arcy Thompson… Bien entendu ces références sont tacites, très loin d’une
officialisation au cœur même des programmes. Plus implicites encore, les
modèles d’inspiration « sonigienne » font irruption dans les manuels
scolaires, de façon parfaitement anonyme mais aussi très significative…
b.
Sur la génétique
Arrêtons-nous donc sur la notion de spécificité
moléculaire ; tantôt implicite, tantôt explicite, son désaveu est
l’événement de loin le plus visible et le plus choquant aux yeux des
enseignants aujourd’hui.
Depuis les années soixante-dix, on enseigne aux
étudiants en biologie les dogmes authentiques de la génétique moléculaire. Les
futurs enseignants y sont formatés dans l’esprit d’une stéréo-spécificité
omnipotente, notion centrale et paradigmatique du réductionnisme génétique,
rayonnant dans toutes les disciplines biologiques :
-
En biochimie ; Toute réaction enzymatique suppose
une parfaite coaptation structurale entre l’enzyme et son substrat. L’enzyme
présente une structure tridimensionnelle fixe, spécifique d’un seul substrat
donné. Le complexe enzyme-substrat renvoie à l’image
« clef-serrure ».
-
En cytogénétique ; Le gène code une enzyme à
fonctionnalité hautement définie. Son expression est contrôlée par des enzymes
inhibitrices ou activatrices reconnaissant spécifiquement la séquence
nucléotidique de ce gène.
-
En immunologie ; La production d’anticorps dirigés
contre tel type d’antigène, reconnu comme n’appartenant pas aux molécules du
« soi », est prédéterminée génétiquement. La spécificité structurale
anticorps-antigène répond au modèle « clef-serrure ».
-
En neurologie ; Un neurone communique avec son
voisin par des neurotransmetteurs. Ces derniers sont captés par des récepteurs
membranaires spécifiques au niveau du neurone post-synaptique, toujours selon
le modèle « clef-serrure ».
-
En endocrinologie ; A une hormone correspond
toujours un récepteur moléculaire spécifique, initiant la réponse cellulaire au
signal hormonal. On retrouve le modèle « clef-serrure ».
-
En physiologie végétale, matière la plus rétive à ces
dogmes, on calque le modèle « clef-serrure » des hormones animales
sur le mode d’action pourtant très polyvalent de l’auxine, des cytokinines, des
gibbérellines… « hormones » végétales.
Le dernier et l’avant dernier
programme regorgeaient de « spécificité » à toutes les échelles.
Voyant dans cette logique une vertu particulièrement pédagogique, les auteurs
des manuels scolaires de l’époque surenchérissaient avec les schémas de
complémentarité structurale entre molécules spécifiques. Toute molécule
organique figurait sous la forme d’une pièce de puzzle coïncidant avec une ou
plusieurs autres pièces. Ainsi des bases complémentaires de l’ADN, des
complexes enzyme-substrat, anticorps-antigène, ou encore
neurotransmetteur-récepteur membranaire. Ce type de schématisation est vite
devenu une routine pour les enseignants en SVT, et l’élève qui savait manier
correctement cette notion obtenait les meilleures notes.
A l’étonnement général, il
n’est quasiment plus question de spécificité dans les nouveaux programmes.
Témoin le chapitre typique d’immunologie en terminale S ; figurant en
bonne place dans le programme précédent, il était question pendant la période
d’élaboration du nouveau programme de le supprimer purement et simplement, au
prétexte que les autres chapitres y prenaient déjà trop de place. Après
consultation des professeurs, majoritairement convaincus de l’utilité de
l’immunologie en terminale, on décida au dernier moment de le sauvegarder… mais
dans quel état ?
L’ancien programme
préconisait un usage quasi-systématique du terme d’« immunité
spécifique », regroupant tous les mécanismes immunitaires liés à la
reconnaissance spécifique des antigènes. On doit aujourd’hui parler
d’« immunité acquise », notion vague, équivoque, surtout destinée à
taire une polémique sans avoir à s’en justifier. Faire référence aux
« caractères acquis » avec une telle insouciance, en immunologie
(« évolution du phénotype immunitaire ») comme en endocrinologie
(« évolution du phénotype sexuel » par exemple), c’est revenir sur
des décennies de martèlement mendélo-morganiste sans pouvoir cacher une
inhabituelle faiblesse théorique !
On passe ainsi
accidentellement d’une notion fortement mécaniste, celle d’immunité spécifique,
à celle d’immunité acquise, ancrée dans une conception générale de l’évolution,
du processus, du mouvement…
De l’ancien au nouveau
chapitre d’immunologie[10]
(fig.9), on passe d’un texte résolument tourné sur les notions de
reconnaissance spécifique et de distinction stricte du « soi » et du
« non-soi », exempt par ailleurs de celles de caractère acquis et
d’influence du milieu, à un texte qui ne fait plus référence à la distinction
« soi / non-soi », très peu à l’immunité spécifique et souvent aux
caractères acquis et à l’environnement… Ce n’est pas une coïncidence, loin de
là. Avançons deux preuves facilement identifiables.
Premièrement, la même
évolution caractérise le chapitre de génétique178 en première
S ; la notion d’« information génétique » n’y est quasiment plus
citée. La métaphore du « programme génétique », vieux réflexe de
biologiste, a totalement disparu. Les références à la stéréo-spécificité se
sont raréfiées de façon significative. En revanche, les notions d’influence du
milieu sur la formation du phénotype est fortement mise en avant ! Elle
était complètement passée sous silence dans l’ancien programme.
Deuxièmement, un article de
correction du programme, publié tardivement au Journal Officiel du 7 août 2002,
supprime sèchement deux des douze références à la notion tabou : « Dans
la partie intitulée I.7. Immunologie : (…) la phrase « les
lymphocytes T cytotoxiques sont aussi des effecteurs de l’immunité
spécifique » est
remplacée par « les lymphocytes T cytotoxiques sont aussi des effecteurs
de l’immunité acquise ». Le titre du paragraphe « les lymphocytes T4,
pivots des réactions immunitaires spécifiques » est remplacé par « les
lymphocytes T4, pivots des réaction immunitaires acquises ». » Ce
nettoyage théorique n’a donc rien de gratuit. On ne trouvera pourtant dans
aucun Journal Officiel de justification ou d’allusion à ces directives.
Dans le document
d’accompagnement aux programmes en revanche, on se montre un peu plus
explicite : « La distinction classique entre immunité non
spécifique et immunité spécifique est obsolète. En effet, de nombreuses
cellules du système immunitaire dites non-spécifiques interagissent avec leur
cible grâce à des récepteurs membranaires qui présentent un certain degré de
spécificité. Les récepteurs des cellules dites spécifiques sont des récepteurs
qui peuvent subir au cours de la réaction immunitaire une modification de leur
structure qui améliore leur capacité de reconnaissance de l’antigène. La
distinction entre immunité à médiation cellulaire et immunité à médiation
humorale, certes classique, est source de confusion dans l’esprit des élèves. »
Pourquoi ne pas l’avouer ; dans l’esprit des professeurs aussi ! Mais
la mauvaise foi revient vite : « Compte tenu du temps imparti à
l’étude de cette partie, des concepts majeurs de l’immunologie ne sont pas
étudiés » Par le plus grand des hasards, ces concepts
« majeurs » aujourd’hui proscrits sont précisément ceux qui décrivent
les mécanismes de l’immunité spécifique !
Dans le chapitre de génétique
en première S, on retrouve la même pudibonderie théorique (fig.9) :
Le document d’accompagnement réaffirme les bases de l’enzymologie
orthodoxe : « La notion de site actif, résultant de la structure
spatiale de la protéine, est précisée : on distingue site de
reconnaissance, en liaison avec la spécificité de substrat, et site
catalytique, en liaison avec la spécificité d’action. », mais dresse
plus loin la liste des notions interdites… ce sont précisément celles qui
permettent d’appréhender la spécificité moléculaire dans le détail :
« L’étude de l’allostérie, l’étude des coenzymes, la discussion sur les
limites de la spécificité des enzymes, la nature des liaisons enzyme-substrat
au niveau du site actif. » C’est avec le même goût du paradoxe que ce
document commente la partie relative à l’expression génétique. Nous sommes loin
des notions exigibles de l’ancien programme : « La notion de gène
morcelé, le contrôle de la transcription [tabou sur les facteurs
spécifiques influençant l’expression génétique], la maturation des ARN [mystère
de la commande génétique hypercomplexe de cette maturation], la maturation
et le devenir des protéines [tabou de la relation unilatérale un gène – une
protéine], la notion d’ARNt [tabou sur les mécanismes du code génétique
lui même], la structure du ribosome » sont désormais
interdites ! Il s’agissait pourtant des découvertes les plus fondamentales
de la génétique moléculaire !… L’auteur du texte s’offre une confession
que le programme officiel ne peut encore se permettre : L’un des
« objectifs [de ce chapitre] est de montrer la complexité des
relations entre génotype et phénotype et de casser l’idée d’un déterminisme
strict entre allèle et phénotype macroscopique ». Luxe bien modeste
finalement ; rien ne permet encore de nettoyer cette confession du mot
« strict » !
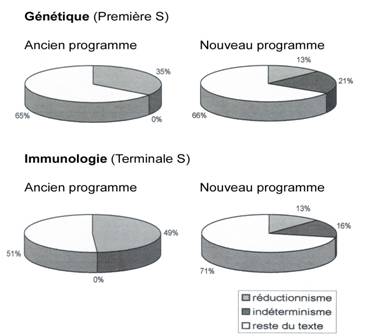
|
|
Réductionnisme |
Indéterminisme |
||||
|
Information gén. |
Programme gén. |
Spécificité |
Environnement |
Milieu |
||
|
Génétique,
ancien programme de IeS. 713 mots |
10
fois |
2
fois |
3
fois |
0 |
0 |
|
|
145
mots : 20% |
94
mots : 13% |
0% |
||||
|
Génétique,
nouveau programme de IeS. 353 mots |
2
fois |
0 |
2
fois |
5
fois |
1
fois |
|
|
29
mots : 8% |
19
mots : 5% |
73
mots : 20,6% |
||||
|
|
Distinction soi / non-soi |
Immunité spécifique |
Environnement |
Acquis |
||
|
Immunologie,
ancien programme de TS. 770 mots |
14
fois |
16
fois |
0 |
0 |
||
|
378
mots : 49% |
0% |
0% |
||||
|
Immunologie,
nouveau progr.
de TS. 1290 mots |
10
fois |
0 |
4
fois |
8
fois |
||
|
162
mots : 12,6% |
198
mots : 15,3% |
|||||
Fig.9 ; Changement
implicite de paradigme du nouveau programme de SVT (Lycée).
Une comparaison des manuels scolaires de l’ancien et du
nouveau programme montre à quel point les auteurs sont désappointés devant
cette schizophrénie théorique (fig.10). Qu’il s’agisse des anticorps ou des
neurotransmetteurs par exemple, les schémas de molécules sont de
plus en plus
fantaisistes. On les classera en trois catégories, qui peuvent
d’ailleurs coexister dans un même manuel ; 1) les schémas typiques dans
lesquels figurent des molécules spécifiques de type « puzzle », 2)
ceux qui sont contraires à ce modèle, figurant des molécules dont les formes
sont volontairement inadaptées deux à deux, 3) ceux enfin, qui évitent pudiquement
les représentations simplistes pour représenter des molécules sans convexité ni
concavité mais censées se coller les unes aux autres sans raison apparente.
Certaines molécules ne se distinguent même plus par leur dessin mais par leur
couleur !
Dans
les années 1980, la première catégorie fait incontestablement l’unanimité. Dans
les années 1990, la troisième catégorie apparaît mais reste fortement
minoritaire, tandis que la deuxième est tellement marginale qu’on la dira
accidentelle. Aujourd’hui, les deuxième et troisième catégories dominent dans
la plupart des manuels.
Manifestement
les manuels scolaires ont beaucoup de peine à suivre l’élan indéterministe
imposé par les nouveaux programmes… Parallèlement au développement marginal des
schémas contraires à la norme de représentation (qu’ils soient maladroits ou
volontaires, c’est à dire destinés à heurter les idées reçues sur la question),
les schémas orthodoxes cèdent en partie la place à des schémas qui évitent de
représenter les molécules au moment précis de la reconnaissance, voire de
représenter ces molécules elles-mêmes ! On ne dira pas pour autant que les
schémas orthodoxes sont rares, bien au contraire (41%). Renversement théorique
non assumé… mais très éloquent ! [voir en annexe la liste des manuels
étudiés et quelques schémas typiques des trois catégories distinguées ici]
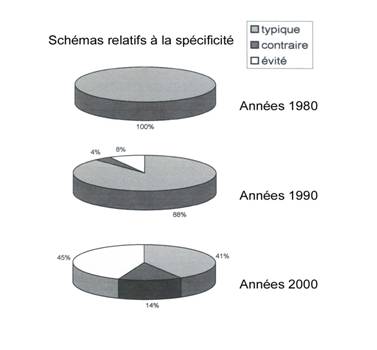
Fig.10 ; Développement
d’une contradiction dans la représentation de la stéréospécificité dans les
nouveaux manuels scolaires.
L’indéterminisme a désormais
le champ libre, profitant d’une crise du réductionnisme pour décréter du même
coup la fin du déterminisme. Cet assaut que nous avons mis en évidence dans le
chapitre 3 doit être l’objet de toute notre attention. Il faut savoir discerner
le juste développement du holisme, à travers la résurrection opportuniste de la
cybernétique (Kupiec, Sonigo, Atlan) et celui de l’indéterminisme qui en reste
inséparable… faute de matérialisme dialectique. Ce développement apparaît dans
les programmes sous son aspect indéterministe, jamais sous son aspect holiste.
Là se tient le blocage que nous voulons mettre en lumière. Rappelons
ceci : la génétique moléculaire réductionniste est née d’une forme de la
cybernétique initialement holiste. Si sa relativisation dans les programmes ne
fait aucun doute aujourd’hui, la cybernétique holiste ne refait pas surface en
tant que telle[11].
En conséquence, le vide
théorique s’épaissit, mettant la biologie officielle dans une situation
instable et contradictoire, mais ponctuellement intéressante ; celle d’une
tentation inavouée, inconsciente, latente du matérialisme dialectique…
C.
Deux pas en arrière : Un darwinisme honteux
a.
Appauvrissement théorique des programmes officiels
Les programmes officiels
explicitent un certain nombre de notions exigibles que les élèves doivent
s’approprier. En matière de génétique, on peut distinguer et compter celles qui
relèvent de la génétique formelle (Mendel), de la cytogénétique (Morgan), de la
génétique moléculaire (Monod), de la génétique des populations (Mayr) et de la
génétique du développement. Rappelons que la « nouvelle synthèse
évo-dévo », stade ultime auto-proclamé de la génétique réductionniste,
interprétant l’embryogenèse et l’évolution à la lumière de ces « gènes
homéotiques » si prometteurs, est au cœur de la tourmente dans la crise
actuelle de la biologie. A l’exception de cette dernière, tous les aspects
historiques de la génétique se présentent aujourd’hui sous une forme
« restreinte », ayant perdu progressivement un certain nombre de
leurs notions-clefs, tout en conservant les plus fondamentales (fig.11).
Des années 80 aux années 90
puis aux années 2000, la génétique formelle perd les notions de facteur
héréditaire, de carte factorielle, de codominance, et même les fameuses lois de
Mendel (leur formulation exacte). La cytogénétique de Morgan perd de nombreux
éléments descriptifs de la biologie cellulaire, gagne entre temps les notions
weismanniennes de lignées somatique et germinale, perd finalement la plupart
des éléments descriptifs fondamentaux pour ne retenir que le strict nécessaire
à l’appui de la génétique formelle. Le mendélo-morganisme est ainsi dilué, mais
conserve son cadre explicatif général [voir liste des notions en annexe].
Evolution du nombre de notions relatives aux différentes branches
historiques de la génétique sur les trois derniers programmes de SVT
(première et terminale S).
Fig.11
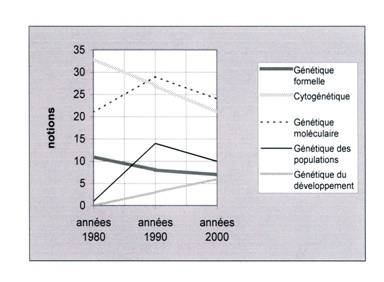
Parallèlement à cette lente
restriction des notions mendélo-morganistes pures, on assiste pendant les
années 90 au développement des notions relatives à la génétique moléculaire
puis à son déclin dans les années 2000, marquant le célèbre retard de
l’enseignement sur la recherche. Remarquons que le programme des années 90
correspond en fait, à travers la multiplication des notions, à leur
renouvellement qualitatif. Les notions les plus mécanistes du programme des
années 80 sont partiellement effacées (mécanismes précis de la synthèse
protéique par exemple), remplacés par les notions moins déterministes du
néo-connexionisme (programme génétique, codage, information génétique,
message). Quelques unes de ces notions seront effacées dans le nouveau
programme, mais on retrouve les notions essentielles du dogme
néo-connexionniste.
La génétique des populations
enfin se développe parallèlement, mais plus lentement : 14 notions dans
les années 90 contre une seule dans les années 80. Les notions se raréfient
déjà dans le nouveau programme avec 10 notions seulement. C’est à l’essor de la
génétique du développement que nous assistons, avec une lenteur plus évidente
encore, entre l’ancien (3 notions) et le nouveau programme (6).
Développement et déclin de
ces disciplines successives n’ont donc pas la même intensité. Le
mendélo-morganisme décroît lentement, comme la génétique moléculaire, tandis
que la génétique du développement, objet pourtant de tous les espoirs, ne se
développe pas avec la vitesse attendue…
En additionnant toutes les
notions de génétique à travers ces trois périodes, la baisse de régime du
renouvellement théorique est très net : on passe d’un total de 66 à 81
notions dans les années 90, puis à 68 notions seulement aujourd’hui !
(fig.12)
Si le développement
qualitatif et quantitatif de l’enseignement de la génétique entre les années 80
et 90 n’a rien d’étonnant, son déclin dans le nouveau programme surprend en
revanche. En effet, dans la même période, une partie de l’enseignement de
géologie prise au hasard, celle qui concerne la convergence lithosphérique,
connaît un développement parfaitement continu : On conviendra que la
géologie est un refuge théorique « neutre » dans le domaine de la
recherche comme dans celui de l’enseignement !
En comparant les listes de
notions exigées par les trois programmes de génétique [voir annexe], on remarquera
qu’il s’agit moins d’une évolution qualitative, portée par des notions moins
nombreuses mais plus complexes, que d’une baisse quantitative quasi-exclusive
des notions enseignées. Deux conclusions s’imposent. 1) Cette baisse est le
signe du malaise des scientifiques face aux échecs de leur modèle théorique
jadis triomphant, 2) le mendélo-morganisme est encore loin du désaveu
total ; les notions les plus fondamentales du modèle, à l’exception de la
stéréo-spécificité que nous avons traitée précédemment, résistent à cette
purge, elle même lente à venir par ailleurs…
Parallèlement à l’élan
indéterministe évident qui marque les nouveaux programmes, la biologie
officielle se trouve contrainte, faute de théorie alternative acceptable, de
faire deux pas en arrière en réaffirmant ses valeurs réductionnistes obsolètes,
mendélo-morganisme d’une part, génétique moléculaire d’autre part.
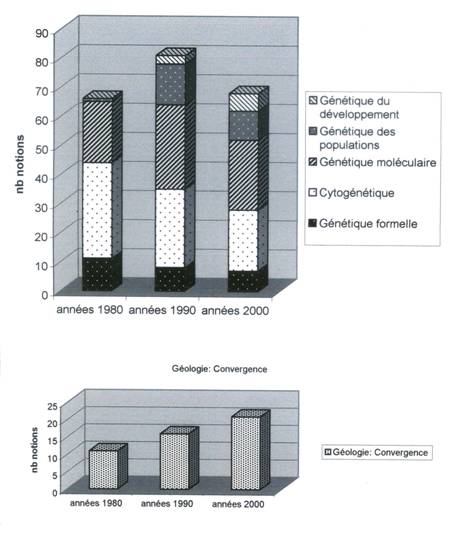
Fig.12 ; Evolution
cumulée du nombre de notions relatives à la génétique et évolution du nombre de
notions relatives à un chapitre idéologiquement neutre de géologie durant la
même période.
b.
Position atypique, instable, décisive
Avant de clore ce chapitre
sur le point le plus important, un détour par l’enseignement de philosophie
s’impose. Les trois derniers programmes ne se sont pas succédés sans heurts
dans cette discipline. Depuis les années 80 jusqu’en 2001, l’avant dernier
texte proposait un thème sur « le vivant », comportant des réflexions
plutôt matérialistes sur l’évolution, sans doute influencées par la cybernétique,
et même des textes de Darwin. En 2001, un nouveau programme fut proposé :
Toujours des éléments d’épistémologie générale … mais plus aucune référence à
la biologie en particulier. Hasard ?
Signalons que Luc Ferry,
dirigeant la rédaction des programmes avant d’accéder au poste de ministre
chiraquien de l’Education Nationale, est l’un des soixante-quinze membres de la
« Fondation Saint-Simon », célèbre et puissant lobby français composé
de journalistes, de grand patrons et d’intellectuels de droite et « de
gauche éclairée ». Pape du néodarwinisme, François Jacob est par ailleurs
le seul biologiste membre de ce Think Tank. Entre les petits fours et
les coupes de champagne, les réflexions idéologiques que nos deux compères
n’ont pas manqué d’échanger ont dû inspirer, en partie au moins, les nouveaux
programmes de lycée.
Contre cette version 2001,
ouvertement politisée, la lutte des professeurs de philosophie a en partie
payé ; deux ans plus tard, un nouveau programme leur est finalement
soumis. Ce dernier est apparemment moins polémique[12],
bien qu’il confirme l’inévitable officialisation d’auteurs réactionnaires tels
que Karl Popper et Annah Arendt.
Trônant parmi les plus
grands, d’Aristote à Marx en passant par Descartes et Kant, nous retrouvons
Ferry se citant lui même (indirectement bien sûr) dans certains manuels. Les
mains salies par la mièvrerie de ce médiocre « philosophe », l’élève
de terminale trouvera plus loin des textes de Jacob, au chapitre concernant les
sciences de la nature… En effet le « vivant », naguère effacé,
réapparaît dans la version 2003. Importance des titres dans les textes
officiels : on passe ici d’un chapitre modestement intitulé « Le
vivant » avant 2001, au très éloquent « La matière, le vivant et
l’esprit » ! On imagine comment y sont traités les couples de notions
exigibles de « cause/fin, contingent/nécessaire, essentiel/accidentel,
formel/matériel, objectif/subjectif »… Bien entendu, rien n’est dit sur le
darwinisme en tant que tel, moins encore sur la génétique. Seule préoccupation
apparente : réconcilier matérialisme et idéalisme !
Rien de bien nouveau donc, si
ce n’est le caractère de plus en plus explicite de l’idéologie dominante. Si la
biologie refait surface dans le programme de philosophie, pour enfoncer le clou
de l’indéterminisme et bannir une fois
pour toutes le
« finalisme »,
rien n’aidera l’élève
à porter un
regard critique sur l’enseignement de SVT. Ce regard critique
apparaît pourtant aujourd’hui plus qu’hier de toute première importance…
Pourquoi le programme de
terminale S en SVT est-il si atypique ? Reprécisons les termes de la
contradiction qui le travaille : 1) Du point de vue strictement théorique,
les auteurs du programme doivent rendre compte de connaissances sur une matière
vivante qui procède dialectiquement, en dressant entre elles des cloisons
« métaphysiques » ; la génétique est la génétique, l’embryologie
est l’embryologie, etc. Telle est la « stratégie » de la science
bourgeoise. Si le courant néopositiviste du XXe siècle cherchait, vainement,
les instruments d’une lutte contre la métaphysique, dans la
« physicalisation » très formaliste de la biologie à l’intérieur des
sciences de la nature, dans la « génétisation » de toutes les
disciplines à l’intérieur de la biologie elle-même, sa forme ultime d’obédience
kuhnienne, enregistrant le reflux de la génétique moléculaire, traduit le grand
retour de la métaphysique en sciences, encourageant les biologistes à une sorte
de néo-vitalisme improbable. 2) Du point de vue pédagogique, une bonne
« progression » se doit de tracer un fil directeur à travers les
chapitres d’une année scolaire, même si ces derniers n’ont apparemment rien de
commun.
Dans l’ancien programme,
cette contradiction est peu marquée. En effet, un matérialisme néopositiviste
fortement teinté de mécanisme fait suite au triomphe de la cybernétique
néo-connexionniste dix ans plus tôt dans le domaine de la recherche. La crise
de la génétique ne fait que poindre et l’introduction au programme officiel
multiplie les allégeances au « programme génétique » et invite au
culte du « mécanisme », qu’il s’agisse de biologie ou de géologie (la
« machine-Terre »). Le fil directeur est donc tout trouvé ; ce
sera l’omnipotence du réductionnisme mécaniste.
Pour les
sciences de la vie : En seconde, le programme comporte la mise en évidence
de différents niveaux d’organisation du vivant (écosystème, organisme, cellule)
et l’étude de l’organisation
fonctionnelle des êtres vivants, sous la dépendance de leur programme génétique et
des facteurs du milieu. En première S, l’enseignement implique la mise en jeu
cohérente des concepts de programme
génétique, de transformation de la matière, de flux d’énergie. Les
acquisitions en chimie permettent d’élucider les mécanismes cellulaires et la manière dont ils
contribuent à assurer l’identité de l’individu et le métabolisme, ainsi que
ceux qui interviennent dans les cycles de matière et le flux d’énergie au niveau des
écosystèmes, et finalement de l’écosystème Terre. En terminale S, l’étude des mécanismes de la
reproduction sexuée, à l’origine de la diversité génétique des individus,
s’appuie sur les connaissances acquises en classe de première à propos de la
nature du matériel génétique et des modalités de son expression, ainsi que de
la relation entre génotype
et phénotype ; l’étude du fonctionnement intégré de l’organisme s’appuie sur
les connaissances relatives à la communication acquises en classe de
seconde ; l’étude de l’évolution biologique s’effectue en liaison avec
celles des milieux et de la géosphère.
Pour les
sciences de
La double
progression, en sciences de la vie d’une part, en sciences de la terre d’autre
part, souligne les interactions biogéologiques : appréhension, en seconde,
de supports proches, limités, accessibles (sol, gisement d’eau) ;
extension surtout dans l’espace, en première, jusqu’au niveau de la planète
avec les cycles
biogéochimiques ; extension, en terminale, plus largement dans le temps et
dans l’espace, avec l’histoire de la terre et l’évolution biologique.
Le programme
comporte cinq parties qui peuvent être abordées dans un ordre différent, à
condition que soit assurée une progression cohérente. Les quatre premières
parties concernent les sciences de la vie. La première, unicité génétique des
individus et polymorphisme des espèces, prolonge l’étude du programme génétique des
êtres vivants réalisée en première. La compréhension de l’unicité des
organismes et du polymorphisme des espèces passe par celle des mécanismes de la
transmission de l’information
génétique et prépare l’étude de l’évolution. La deuxième partie, mécanismes de l’immunité,
permet de réinvestir et d’élargir les acquis précédents et ceux de première sur
le programme génétique.
Elle apporte en particulier une illustration de son expression polymorphe et
une explication du maintien de l’intégrité du soi au niveau moléculaire. La
troisième fonde la compréhension de quelques aspects du fonctionnement des
centre nerveux sur l’étude des mécanismes à la base des propriétés des neurones et réseaux
neuroniques. Elle s’appuie sur l’étude de la motricité somatique. La quatrième
porte sur le fonctionnement d’un système de régulation, à l’échelle de l’organisme, dans le cas
des taux d’hormones sexuelles mâles et femelles.
La cinquième
partie associe sciences de la terre et de la vie dans un même thème général :
histoire géologique et évolution biologique. Ce thème est l’aboutissement
logique des études qui, en classe de seconde, situent notre planète au sein du
système solaire et, en classe de première, abordent les manifestations internes
et externes de la dynamique terrestre. Il permet d’esquisser la fresque des
grandes étapes de l’histoire de la terre, grâce à l’étude des modifications de
sa surface et au constat des transformations de la biosphère.
BO spécial n°6 du 9 juin 1994
Il en va tout autrement du
nouveau programme. Un fil directeur fédérera t-il des chapitres aussi
différents que l’évolution biologique, la géologie, l’immunologie et
l’endocrinologie sans mécanisme réductionniste ?
Par un heureux
« hasard », c’est dans ce contexte que nous retrouvons la
dialectique. Attention : L’introduction au nouveau programme de TS est
loin d’une apologie du matérialisme dialectique ! Elle est précisément
l’illustration concrète de l’impossibilité/nécessité d’une conversion de la
science bourgeoise au matérialisme dialectique. L’illustration du caractère à
la fois salutaire et délétère de la position actuelle
anti-réductionniste/anti-déterministe. La voici :
Notre
planète, tant en ce qui concerne la géosphère que la biosphère présente deux propriétés d’apparence
contradictoire : stabilité et variabilité. Cette contradiction se résout
par la prise en compte de la dimension
temporelle. L’un des objectifs du programme de la classe de terminale
est de fournir un modèle dynamique de
Le monde vivant
présente une unité structurale et fonctionnelle mais aussi une très grande
diversité ; cette diversité lui permet de se maintenir globalement au cours du temps et de
s’étendre dans l’espace. Ainsi, la stabilité de la biosphère s’accompagne de la variabilité des espèces
(évolution), la stabilité
de l’espèce s’accompagne de la variabilité des individus (procréation, génétique), la stabilité de l’individu
s’accompagne de la variabilité
de certains de ses constituants (par exemple le système immunitaire).
Comprendre l’évolution
biologique et géologique de la planète requiert la capacité d’identifier des
moments remarquables dans l’histoire de la terre, de les ordonner, d’évaluer
leur âge et de mesurer les durées
qui les séparent.
Approche du temps en biologie et en
géologie : L’objectif de cette [présentation] est d’introduire
le programme, de lui donner du sens, d’en faire percevoir le fil directeur et
la logique. Il s’agit plus précisément de conduire les élèves à s’interroger
sur les différentes échelles de temps utilisées pour comprendre l’évolution conjointe de la planète et de la
biosphère. Cette introduction s’appuie sur le perception empirique du temps qu’ont les élèves.
Elle ne constitue en aucun cas une liste de contenus ou de notions exigibles au
baccalauréat. Toutefois, si une notion utilisée dans l’introduction est reprise
dans une autre partie du programme, elle pourra alors faire l’objet de
questions à l’examen, ces questions se cantonnant exclusivement aux contenus et
respectant les limites de la partie du programme correspondante. Les
indications ci-dessous ne sont que des propositions.
Comment la
planète actuelle s’est-elle construite au cours du temps ? Son fonctionnement a t-il
toujours été conforme à l’actuel ou s’est-il modifié au cours du temps ? Quels sont
les évènements majeurs qui jalonnent cette histoire ? Quand se sont-ils
produits ? Comment peut-on les dater ? Comment peut-on apprécier leur
durée ? Sur
quels critères notamment temporel,
peut-on définir la stabilité ou la variabilité d’un individu, d’une chaîne de
montagne, d’une molécule, d’une espèce, d’un domaine océanique ? Quelles
sont les durées
caractéristiques d’existence d’un individu, d’une chaîne de montagne ? Les
modifications de la planète et de ses habitants sont-elles continues ou discontinues ?
Méthodes et
supports envisageables : Repérer sur une frise du temps les grands évènements déjà abordés au
cours des classes précédentes en sciences de la vie et de la terre. Par une
recherche documentaire, faire construire une frise du temps en y plaçant les évènements
couramment évoqués dans la presse scientifique. Parmi les évènements clés
intéressants, on peut citer sans les développer, les exemples suivants :
La formation de la terre, l’apparition de la vie, l’apparition d’une atmosphère
oxydante, la mise en place de la tectonique des plaques, l’apparition du
premier eucaryote, etc.
Faire
classer les grands évènements biologiques et géologiques selon leur durée. Faire naître un
questionnement sur le mode d’appréciation de la durée des phénomènes. Discuter sur un exemple de
la continuité ou
discontinuité d’un phénomène selon l’échelle de temps utilisée pour le décrire. Discuter
sur un exemple de la stabilité ou de la variabilité d’un objet, d’un mécanisme,
etc. en fonction de la durée
de son observation. Parmi les « objets » et « mécanismes »
dont on peut apprécier la durée :
la planète Terre, une chaîne de montagne, une période glaciaire, une espèce, un
individu, une cellule, etc.
BO hors série n°5 du 30 août 2001
Premier constat : Plus
aucune trace des notions de programme génétique ou d’information génétique. Le
terme de « mécanisme » lui même ne revient plus qu’en tant que
vieille habitude encore mal combattue, quand il n’est pas tout simplement entre
guillemets. En revanche, un consensus s’établit autour des notions de
« temps », de « durée », d’« évolution »… voilà
donc, très explicitement, le nouveau fil directeur !
|
Introduction
au programme de TS |
Avant |
Après |
|
|
Programme génétique |
Mécanique |
4 |
0 |
|
Information génétique |
1 |
0 |
|
|
Mécanisme |
6 |
2 |
|
|
Machine |
1 |
0 |
|
|
Evolution |
Dialectique |
5 |
4 |
|
Temps |
2 |
10 |
|
|
Continuité/discontinuité |
0 |
2 |
|
|
Durée |
0 |
7 |
|
|
TOTAL |
12
mécanique 7
dialectique |
2
mécanique 23
dialectique |
|
Fig.13 ; Le rejet du
mécanisme provoque l’apparition de la dialectique dans les textes officiels.
Les deux premières phrases du
présent texte sont étonnement claires. Elles invitent à une approche presque
ouvertement dialectique de la matière, qu’elle soit vivante ou non ! La
métaphysique vitaliste semble donc provisoirement renvoyée au placard.
Considérant la matière dans son évolution, cette introduction qui jette la
trame du programme annuel fait plus encore ; elle omet du même coup et
sans justification ce mécanisme qui traversait les anciens programmes avec le
ton de l’évidence.
Si certains formateurs IUFM
font du « temps » le pivot central d’une progression idéale, d’autres
lui préfèrent carrément le concept central de
« contradiction » ! Comme entraînés contre leur gré par cet élan
dialectique complètement inédit, les auteurs du programme passent même de la
notion de contradiction à celle de continuité/discontinuité ! Arrive t-on
au saut qualitatif ? Bien sûr que non. Mais rien n’empêche plus un
enseignant marxiste de l’enseigner légitimement ! (fig.13)
Modérons toutefois notre enthousiasme…
Les « deux propriétés » que « présentent notre planète, tant en
ce qui concerne la géosphère que la biosphère » ne sont
qu’« apparemment contradictoires », même si le texte qui suit atteste
la réalité de cette contradiction. De plus, on affirme naïvement que
« cette contradiction se résout par la prise en compte de la dimension
temporelle » : Cette prise en compte ne peut être que saluée, même
si, rappelons-le, elle est incroyablement tardive, mais la dimension temporelle
ne saurait être la clef d’une réelle compréhension des évolutions géologiques
et biologiques. On cherche à expliquer une contradiction antagoniste évidente
par l’évolution elle même, c’est à dire son effet. On cherche en somme à
élucider un miracle… alors que cette contradiction est précisément le moteur,
la cause nécessaire de l’évolution !
Ainsi renversée, la
démonstration n’a aucune chance d’appeler un jour le secours du matérialisme
dialectique, moins encore d’aboutir à de nouvelles connaissances réelles… Et
nous retrouvons la vieille « perception empirique » qui fera primer
des aspects secondaires, comme la distinction des échelles de temps, sur
l’aspect principal du problème…
Prolongeant la recherche
officielle, l’enseignement est ainsi un puissant révélateur des blocages de
l’idéologie dominante. Par delà la contradiction
anti-déterminisme/anti-réductionnisme, qui explique les tentations dialectiques
plus ou moins explicites des Tort, Jacob, Gros, Gould, Kupiec et Sonigo, etc.,
les aveux implicites des auteurs du programme pourraient à la rigueur
convaincre aujourd’hui des esprits jadis réfractaires au marxisme, mais sont
surtout pour nous, par leur caractère implicite même, autant de preuves de ces
« honteuses » tentations et de leur échec annoncé.