Chapitre 6 : Hérédités et pression désorganisatrice
Théorie dialectique
de l’hérédité
« On peut y arriver [à la conception dialectique de
F.Engels (Dialectique de
Si l’approche dialectique donne à l’histoire de la vie un cadre
descriptif cohérent qui rend à l’évolution darwinienne sa nécessité
fondamentale, bien au delà du simple mécanisme hasard/sélection, elle consiste
à effectuer un renversement théorique de la génétique instructionniste, c’est à
dire à bâtir partant d’elle les instruments de sa propre négation dialectique
(A).
En nous conformant à ce que préconise Engels, c’est à dire en portant la
« conscience des lois dialectiques au devant du caractère dialectique de
ces lois », nous nous sommes départis de l’écueil empiriste consistant à
juxtaposer des concepts à valeur descriptive sans tenter d’en identifier les
interactions objectives. Nous en tirerons ici les conséquences :
Authentifier l’existence d’une contradiction objective opposant hasard et
sélection naturelle, inscrire le moteur de l’évolution non dans cette
« opposition » à caractère métaphysique, c’est à dire exclusivement
dans l’un ou l’autre de ses termes, mais dans le dépassement dialectique de
leur contradiction dynamique (B).
A. Les bases génétiques d’ une théorie dialectique de l’hérédité
C’est de la notion de gène telle qu’elle fut définie par Crick et ses
coreligionnaires, que naît la génétique moléculaire dans les années cinquante,
en même temps qu’un renouvellement de la conception vitaliste selon laquelle la
vie au sens large tient moins aux lois matérialistes de la chimie qu’à celles
de l’« information », de la « stéréospécificité », du
« programme ».
Dans l’option (la nôtre) qui consiste à poser pour objectif malgré leur
interprétation les résultats de nos chers praticiens, nous avons vu à quel
point il est facile de la couvrir d’une dangereuse téléologie ; Cette
molécule « est produite pour », cet organe « sert à », ce
gène « code » tel caractère… Que de dérives naissent de telles
métaphores ! Résumons les.
Problème des finalistes : Si chacun de nos organes concourre à un
rôle précis dans l’organisme (stocker les sucres pour le foie, assurer
l’oxygénation du sang pour les poumons) à l’instar des pièces d’une machine ou
d’un robot, à quoi sert donc la machine elle-même ?
Nous avons le choix ; 1) L’organisme est le véhicule de l’âme, 2)
l’organisme est le véhicule des gènes qui l’ont façonné (véhicule sans lequel
ils ne sauraient subir la sélection naturelle !), 3) l’organisme est
l’« organe » d’une collectivité d’organismes à un niveau supérieur,
4) l’organisme est structuré de telle façon qu’il conserve sa structure… Nous
voyons à quel point cette dernière solution, qui n’est pas forcément en
contradiction avec la troisième, revêt une « téléologie » spéciale et
apparaît pour les lecteurs peu familiers de la dialectique, sous la forme d’une
audacieuse tautologie ! Mais la vulgate darwinienne n’en est-elle pas une
autre ?
Ni la population, ni l’organisme, ni même la cellule ne sont donc le
« véhicule » des gènes, pas plus que ceux-ci ne sont les pages d’un
« Livre de Vie ». Seule certitude ; un gène est une portion
d’ADN qui contrôle la production d’un ou de plusieurs type-s de protéine-s au
niveau cellulaire. Comme les cellules fonctionnent ensemble dans un organisme
pluricellulaire, il n’est pas étonnant qu’un défaut génétique ponctuel ait des
répercutions nombreuses et variées sur le phénotype de cet organisme. De là à
affirmer que tous les caractères visibles et invisibles de notre organisme
correspondent chacun à un gène précis, le fossé est vite franchi…
Remplaçons l’ADN par un cobaye dans le laboratoire du généticien et
remplaçons le matériel de génie génétique par un simple bistouri : Que le
cobaye meure après une ablation de ses reins et notre praticien dira que le
rein est l’organe qui sert à éviter la mort. Que le cobaye fasse une
grosse dépression une fois castré, il dira que le testicule est l’organe du
bon moral !
Mais c’est bien plus qu’une question de méthode. La définition du gène a
beaucoup évolué depuis un siècle. Que chaque caractère phénotypique requiert le
concours de nombreux gènes, que chaque gène participe à l’élaboration de
nombreux[1]
caractères phénotypiques, cela ne fait aujourd’hui plus de doute. Il semble
bien difficile de définir individuellement le gène. C’est du reste le
même problème au niveau de l’organisme : Chaque organe concourre à de
nombreuses fonctions, sans rapports entre elles, chaque fonction physiologique
est assurée par la participation de nombreux organes. C’est sous l’angle d’une
observation à plusieurs niveaux, et surtout dans les interactions
qu’entretiennent ces niveaux entre eux, qu’on peut remettre l’ADN à sa juste
place.
Une conclusion provisoire s’impose : Nous ne nions pas l’existence
des gènes, ni la place qu’ils occupent dans les processus cellulaires.
Cependant la place faite aux gènes dans la théorie mendélienne de l’hérédité
mérite sérieusement d’être attaquée, au sens où elle constitue une théorie au
plus haut point idéaliste : Pour nous, au niveau de leurs processus,
hérédité cellulaire et hérédité des organismes pluricellulaires seraient
superposables et non confondues. Ce sont des processus reproductibles, non des
informations abstraites transmissibles. Aux processus héréditaires de la
cellule, au moins partiellement relatifs aux gènes, s’ajoutent des processus
héréditaires propres au niveau pluricellulaire.
Un exemple marquant le caractère incomplet d’une théorie génétique de
l’hérédité pour les organismes pluricellulaires : L’expérience pionnière
du clonage reproductif chez les mammifères avec la brebis Dolly récemment
mérite un intérêt particulier. Cette brebis est née d’une cellule-œuf ayant
shunté l’étape de la reproduction sexuée (on a ôté à un ovule ses gènes
haploïdes issus d’une méiose, pour les remplacer par le génome intégral
diploïde de la « mère », c’est à dire par le noyau d’une de ses
cellules somatiques).
Premièrement, ce type d’expérience relève plus d’une
« cuisine » empirique de laboratoire que d’un réel exploit du génie
génétique dérivant de sa théorie fondamentale. Elle devait répondre à des
questions théoriques relatives au rôle du noyau (donc en partie des gènes) sur
le développement embryonnaire et l’acquisition du phénotype. A ce titre, les
auteurs ont constaté contre toute attente, que du noyau et du cytoplasme, c’est
semble t-il ce dernier qui s’avère le plus déterminant ! Le rôle du noyau
est d’autant plus relatif que l’implantation dans la cellule œuf d’un noyau de
cellule somatique cancéreuse produit un organisme parfaitement sain…
Deuxièmement, contre toute attente, la brebis a vieilli précocement,
elle est morte prématurément, malgré les soins extrêmes dont elle a bénéficié.
La méiose consistant fondamentalement en un
brassage génique favorisant le polymorphisme au niveau du groupe
spécifique, elle aurait donc une vertu supplémentaire au niveau de
l’individu. Vertu dont Dolly n’a pas pu profiter. Cette vertu mystérieuse mais
bien réelle 1) intéresse directement l’hérédité pluricellulaire et non
exclusivement l’hérédité cellulaire (la cellule-œuf s’est développée
normalement), 2) concerne à travers cette hérédité non pas la nature de chaque
gène mais l’état du noyau dans sa
totalité, 3) se rapporte à l’histoire de ce noyau (« vieux » noyau
somatique contre « nouveau » noyau issu d’une caryogamie dont la
structure génique serait pourtant la même –diploïde-) plutôt qu’à la structure
décrétée fixe de ses gènes.
Troisièmement, cet échec relatif indique d’une part que la
reproductibilité de la forme pluricellulaire organisée passe effectivement par
des processus nucléaires, donc relatifs aux gènes, d’autre part qu’une
adéquation noyau-cytoplasme (à élucider) est nécessaire à la finalisation de
cette reproduction. Si on considère l’hérédité comme la reproduction d’une
structure fixe (la brebis sous sa forme adulte par exemple), thèse
idéaliste-essentialiste, l’expérience est réussie dans la cas de Dolly. Si on
la considère comme la reproduction d’une ontogenèse complète, c’est à dire d’un
processus fécondation, embryogenèse, développement, sénescence, mort,
thèse matérialiste ; c’est un demi-échec[2].
Trois remarques…
1)
Considéré comme la reproduction
d’une ontogenèse, l’hérédité se rapporte au noyau mais aussi au cytoplasme
(surtout animé par des activités protéiques) : L’expérience consistant à
injecter le « jeune » noyau d’une cellule-œuf dans le vieux
cytoplasme d’une cellule somatique, non armée comme l’est la cellule-œuf
(réserves, forme, …) pour effectuer une embryogenèse, est condamnée à échouer.
Pourtant le clonage thérapeutique consistant à produire in vitro un organe ou
un tissu (reproduction ontogénique partielle) à partir d’une cellule somatique
correspondante isolée donne des résultats satisfaisants.
2)
Le même constat peut être tiré de
l’observation du cycle de développement de nombreux végétaux primitifs par
exemple, chez qui reproduction asexuée (sporulation, bouturage naturel, …) et
reproduction sexuée coexistent. On considère en biologie, bien qu’aucune
explication n’en soit fournie, qu’une telle espèce, en ne pratiquant plus que
la reproduction asexuée (les spores sont des cellules « non-rénovées »
par la méiose) dépérirait en quelques générations.
3)
Toujours en ce qui concerne la
biologie végétale, domaine malheureusement négligé par les néodarwinien
weismannien (et pour cause, puisqu’à la différence des animaux, les végétaux ne
présentent rien qui ressemble à une lignée germinale), la polyploïdie[3]
d’une plante modifie très fortement ses caractères et son hérédité (justifiant
l’apparition artificielle d’une nouvelle espèce). La polyploïdie ne modifie
pourtant en rien l’état qualitatif des gènes de la cellule, elle ne modifie le
noyau que « quantitativement ».
Lors de la session de l’Académie Lénine des sciences agricoles d’Union
Soviétique, où s’affrontaient les partisans de l’orientation mitchourinienne et
les partisans de l’orientation weismannienne, l’académicien S.Démidov
affirmait, en référence à Lénine et Staline, que les contradictions entre deux
courants doivent être résolus non pas par la conciliation et la recherche d’une
troisième voie intermédiaire, mais par une lutte de principe, aiguë et
déclarée.
On constatera aisément, au fil de l’exposition d’une théorie de
l’hérédité en accord avec les conclusions du chapitre précédent, qu’elle paraît
« conciliatrice », s’accommodant des notions de gène, de soma et de
germen, tout en étant objectivement en franche opposition avec la théorie
génétique dans ses principes, c’est à dire dans l’usage qui est fait de ces
notions. Aucun désaccord de principe donc, avec des réflexions matérialistes
telles que celle de Lyssenko :
« Les
expériences sur l’hybridation végétative montrent indiscutablement que toutes
les particules du corps vivant, même les substances plastiques, même les sucs
qu’échangent greffon et porte-greffe, possèdent des qualités héréditaires. Cela
diminue t-il le rôle des chromosomes ? Pas du tout !
L’hérédité se
transmet-elle par des chromosomes au cours du processus sexuel ? Oui, bien
entendu ! Nous reconnaissons l’existence des chromosomes, nous ne la nions
pas. Ce que nous ne reconnaissons pas, c’est la théorie chromosomique de l’hérédité,
c’est le mendélo-morganisme. »
T.Lyssenko, Xe séance de la session des sc. Agricoles de l’URSS, 1948.
a. Premier niveau : L’hérédité cellulaire
Il s’agit en particulier d’examiner à partir de nos thèses le dogme
central de la biologie moléculaire, dogme affirmant que si le gène détermine
sa protéine, la protéine ne peut elle même déterminer son gène. Remarque
préliminaire : Soit nous nions les mécanismes relatifs à la synthèse
protéique tels qu’ils sont exposés par la génétique moléculaire, et la question
du dogme central ne vaut plus d’être posée, soit nous les admettons dans un
cadre qui est celui des mécanismes conservateurs, et l’unidirectionnalité
gène-protéine ne sera plus un dogme idéaliste mais un processus résultant de
l’histoire de la vie et par suite, un moteur de sa propre négation.
« La vie est
le mode d’existence des corps albuminoïdes [protéines],
(…) ce mode d’existence consiste essentiellement dans un perpétuel
auto-renouvellement des éléments chimiques dont ces corps se composent. »
Engels (Dialectique de
Ce sont en effet les protéines qui, par leurs propriétés
physico-chimiques variées, assurent tous les processus cellulaires (enzymes et
protéines de structure). A ces protéines s’ajoute une autre catégorie chimique
dont la vocation est de participer à cet « auto-renouvellement » des
protéines, les acides nucléiques (ADN et ARN). Considérons que les gènes ont
pour vocation principale de renouveler pendant la vie de la cellule, toutes ses
protéines fonctionnelles à mesure qu’elles dégénèrent (mouvement dialectique),
pour vocation secondaire, collatérale, d’être à la base de l’hérédité
cellulaire (mitose). Voyons donc ce qui se passe au cours de la vie
d’une cellule.
Toute la génétique moléculaire repose sur une dualité acides nucléiques
– protéines. Les uns sont inertes, impénétrables, maîtres, les autres sont
actives, invariables et esclaves (déterminées). Nous considérerons au contraire
que ce dualisme n’est qu’un fractionnement historique[4]
(ayant engendré au début de l’histoire de la vie les propriétés conservatrices
de la cellule), que gènes et protéines sont tous variables, à des degrés
différents, qu’ils entretiennent des relations mutuelles dans le cadre de leurs renouvellements respectifs.
En dialecticiens, nous associerons conceptuellement des mécanismes sur
lesquels les généticiens idéalistes se penchent toujours isolément – à l’abri
de toute tentation de péché de synthèse et en sacralisant en chacun les mystères
vitalistes – sous un angle où leur « non problématicité » peut
apparaître.
Premièrement les gènes se tiennent
en amont des processus de renouvellement des protéines constitutives de la
cellule (synthèse protéique). Nous dirons qu’ils contrôlent la synthèse de ces
protéines plutôt qu’ils les déterminent (nous reviendrons sur cette remarque).
Ils en contrôlent la qualité (types de protéines) mais aussi leur quantité
(proportions relatives de ces protéines).
Deuxièmement les protéines assurent partiellement le renouvellement des
gènes eux-mêmes, suivant les processus déjà évoqués au chapitre
précédent : Réplication (dédoublement de l’ADN), transcription (production
d’ARN et d’amorces), duplication (de certains gènes, selon les besoins
ponctuels de la cellule), réparation (systèmes SOS – SRM).
Le renouvellement « à l’identique » des molécules
constitutives de la cellule est réalisé sur la base d’une interaction acides
nucléiques–protéines. Chacune des deux catégories ne peut se renouveler
(propriété de la matière vivante) sans l’autre. Fondée sur les mêmes propriétés
physico-chimiques, l’hérédité cellulaire (conservation des caractéristiques
cellulaires de génération en génération) est indissociable des processus de
renouvellement moléculaire (au cours de la vie d’une cellule). Ces derniers
constituent les bases de la propriété conservatrice de toute matière vivante.
Rappelons notre proposition[5]
de départ : Ce qui distingue la matière vivante de la matière non-vivante,
c’est la tendance qu’elle manifeste à conserver sa structure dans le temps. Si
le processus conservateur de la matière vivante provient à l’origine du
fractionnement qualitatif des molécules fonctionnelles et auto-réplicatives
(fractionnement qui donne lieu au dogme
central), n’oublions pas que les molécules auto-réplicatives (acides
nucléiques) sont encore douées de certaines
activités reconnues (la plus
connue chez les cellules actuelles, est celle des ARN ribosomaux) et que les molécules
actives (protéines) peuvent présenter
une activité « auto-catalytique »[6]
voire carrément auto-réplicative[7].
Bien à l’abri de ces considérations, la génétique moléculaire décrit ainsi la
synthèse protéique :
►
Transcription du gène (ADN) en plusieurs ARN pré-messagers identiques grâce à
une polymérase (protéine) dont on ne sait pas bien pourquoi elle adhère en
amont (région promotrice) de ce gène plutôt qu’en amont d’un autre dans le
noyau, chaque gène ayant sa région promotrice particulière et nos savants ne
connaissant qu’un à trois types de polymérases seulement.
►
Maturation des ARN : Des enzymes (protéines) découpent des fragments
(introns) et réassemblent bord à bord les séquences restantes (à traduire) –les
exons-. Comme tous les introns de tous les gènes sont différents et qu’on
considère chaque type d’enzyme spécifique de son substrat, il faut donc plus de
sortes d’enzymes (quelle est leur origine ?). Que de gènes dans le
noyau !
►
Traduction des ARN messagers en polypeptides (chaînes libres d’acides aminés)
grâce à un énorme complexe enzymatique, le ribosome, composé essentiellement
d’ARN ribosomaux (ARNr). Il faut ajouter que chaque acide aminé ajouté au
polypeptide en cours de synthèse est amené par un ARN de transfert (ARNt),
chaque type d’ARNt correspondant à un type d’acide aminé. L’accrochage des
acides aminés sur les ARNt, préalable à cette opération, est réalisé par de
nombreuses enzymes (elles mêmes déterminées par des gènes !).
► Le
polypeptide n’est encore qu’une chaîne inactive dont seule la combinaison
linéaire d’acides aminés est spécifique ; il subit alors une maturation
complexe destinée à lui faire acquérir la forme finale qui lui conférera une
activité catalytique spécifique. Cette maturation est réalisée au moyen
d’enzymes spécifiques de ces polypeptides, elles mêmes déterminées par des
gènes… Que de gènes, que de gènes !!
Leur nombre total ? On l’estimait chez l’Homme à plusieurs
centaines de milliers il y a quinze ans. Le centre national de séquençage (Génoscope)
donnait timidement il y a trois ans une estimation de seulement 30 000 gènes.
Il en donne finalement, une fois le long travail de séquençage achevé (avril
2003), 25 000 (c’est à dire un nombre proche de celui de nombreuses espèces
unicellulaires elles-mêmes ! Ajoutons, et on le sait depuis longtemps,
qu’en ce qui concerne la quantité d’ADN nucléaire, certains vertébrés primitifs
–des batraciens comme Necturus et Amphiuma par exemple- ont un
génome quantitativement plus important que celui de l’Homme). Quelques
informations intéresseront peut être les gardiens du dogme…
- C’est par complémentarité directe qu’une séquence d’ADN (gène) produit
un ARN pré-messager, la polymérase (protéine) se contente de localiser, d’orienter,
d’ouvrir le gène à transcrire : L’ADN a une fonction catalytique
spontanée. De plus le ribosome traduisant l’ARNm en
polypeptide doit
ses activités
enzymatiques aux seuls ARNr (les petites protéines ribosomales se tiennent en
périphérie du ribosome et non dans son site actif). Plus que de détenir
passivement les plans de construction des protéines, ce sont bien les acides
nucléiques qui président à la production de celles-ci.
- Le phénomène d’épissage alternatif des ARN prémessagers complique
considérablement la tâche des séquenceurs actuels du génome humain. Connu
depuis plus de vingt ans, il s’avère
plus important que prévu chez les
vertébrés eux mêmes… L’épissage alternatif conduit à produire selon les cas, à
partir d’un type donné d’ARN pré-messager (donc d’un gène) et selon les introns
qui sont excisés plusieurs sortes d’ARNm
donc plusieurs types différents de protéines !!
- De nombreux gènes contrôlent la production de protéines appelées
facteurs de transcription (dont l’affinité
pour telle ou
telle région promotrice
de l’ADN dirigerait
dans l’espace et dans
le temps les expressions génétiques[8]).
Problème : On recense fort peu de types de facteurs de transcription (une
dizaine) au regard du nombre de gènes différents dont il faut coordonner
sélectivement les expressions. De plus, chaque facteur de transcription n’est
spécifique que d’une séquence de trois à six nucléotides sur l’ADN, c’est
étonnement peu pour discriminer 25 000 gènes aux promoteurs différents… Cependant
la colinéarité de certains gènes étonne plus encore les chercheurs. Comment
expliquer par exemple que les gènes des ε, γ, δ globines, qui
apparaissent successivement dans le sang du fœtus au cours de sa croissance, se
suivent dans cet ordre sur le même tronçon chromosomique ? Comment
expliquer de même la succession des « gènes du bras », « de
l’avant-bras » et « de la main » sur un autre
tronçon ? L’effet de position dans le noyau semble être fondamental dans
la régulation de l’expression génétique, même s’il est complètement hors champs
dans la génétique instructionniste. Il existerait des séquences d’ADN
« tour de contrôle » à proximité des gènes à contrôler (Duboule, Nature
n°420). Ainsi les gènes ne sont pas si inertes : Ils interagissent… On
peut donc apparemment appliquer la « loi de corrélation des organes »
(Cuvier) aux gènes eux-mêmes mais en développant des thèses tout à fait
opposées à celles du fixisme !
- Le cas du prion cité plus haut laisse supposer que les protéines sont
elles mêmes douées de propriétés auto-catalytiques. Des polypeptides immatures
adoptent spontanément des formes qui ne sont pas celles qu’ils conservent après
maturation. Il est admis qu’un polypeptide peut se replier de plusieurs façons
possibles, statistiquement et en fonction de la température, etc. Pourquoi ces
formes « immatures » n’auraient-elles pas des activités catalytiques
propres dans la cybernétique cellulaire ? L’intrication de ces activités
entre polypeptides, entre enzymes, contribuant à leurs maturations respectives,
peut alors être telle que 1) l’ensemble des synthèses protéiques soit partiellement
détachée des processus d’expression génétique, contre la théorie réductionniste
instructionniste : La production d’une enzyme dépend d’une production
d’ARNm et d’activités transversales d’autres protéines, immatures et/ou
matures. 2) Chaque protéine immature (de structure immature active)
puisse donner, en fonction des activités transversales, plusieurs types
d’enzymes différentes (autres structures, autres activités). Quelle économie de
gènes ! 3) La profonde interdépendance entre toutes ces chaînes de
production de protéines fonctionnelles valide la « loi de corrélation des
organes » appliquée à l’ultra structure moléculaire de la cellule, garant
d’une auto conservation d’ensemble (équilibre dynamique malgré les variations
locales) plutôt que d’un déterminisme strict et applicable a priori en
tout point.
Manifestement, d’un point de vue matérialiste dialectique, la théorie de
l’hérédité cellulaire repose secondairement sur l’interdépendance de deux
contradictions antagonistes, elles mêmes fondées sur un fractionnement initial
d’une catégorie moléculaire, l’ARN, douée à la fois des deux propriétés
conservatrices que sont 1) l’activité catalytique (dont par exemple celle de
construire à partir
de précurseurs naturels ses
propres sous-unités constitutives[9]),
2) l’activité auto-réplicative issue d’une complémentarité spontanée entre deux
nucléotides et fondée sur la polymérisation d’un nouvel ARN sur un ancien ARN
servant de matrice linéaire, en deux catégories qualitativement
distinctes : l’ADN (propriété 2), les protéines (propriété 1).
Première
contradiction antagoniste
La propriété auto-réplicative des acides nucléiques n’est pas
« acquise pour » renouveler le stock d’acides nucléiques en
destruction permanente et naturelle. Elle est inhérente à la structure même de
ces acides nucléiques (complémentarité deux à deux des nucléotides) de sorte
qu’il n’existe a priori aucun couplage naturel entre la reproduction moléculaire
des acides nucléiques et une mystérieuse nécessité de conservation structurale.
Ainsi leur propriété auto-réplicative ne « répond » pas à leur
destruction naturelle (pas de téléologie), elle l’« anticipe »
fortuitement. Initialement les acides nucléiques se multiplient plutôt qu’ils
se renouvellent. Dès lors l’équilibre stabilisant le stock d’acides nucléiques
est fortement dépendant des conditions du milieu, susceptible en variant
d’accélérer ou de freiner l’auto-réplication. La conservation est qualitative
(la structure moléculaire continue d’être représentée dans le temps), elle
n’est pas quantitative : Si le milieu freine la destruction des molécules
(milieu favorable) il favorise la conservation quantitative du stock
moléculaire. Cette conservation de molécules auto-réplicatives augmentera leur
quantité (non conservation quantitative). Bel exemple de contradiction
antagoniste : Au niveau moléculaire, la « matière vivante » tend
donc à se développer parce qu’elle se conserve !
Deuxième
contradiction antagoniste
La matière vivante se conserve parce qu’elle reproduit ses propres
matériaux. Cette reproduction se réalise sur deux niveaux. Le premier est celui
des nucléotides constitutifs eux mêmes (production qui dépend de la quantité
disponible de précurseurs dans le milieu, de la qualité et des proportions
relatives des moyens enzymatiques de production dans la cellule), le deuxième
est celui de la polymérisation de ces nucléotides en acides nucléiques sur la
base d’un acide nucléique polymérisé préexistant.
Une variation quantitative des stocks moléculaires contribuant à ces
deux niveaux affecte donc directement la capacité qu’a la matière vivante de se
conserver (sans remettre en cause la propriété auto-réplicative des acides
nucléiques).
Dans le « monde à ARN » primitif, la cellule ne peut se
conserver qu’au moyen de plusieurs types de « ribozymes » (ARN)
assurant les activités des niveaux 1 et 2. Ces ribozymes, condamnés par leurs
places respectives dans la
cybernétique cellulaire à
rester distincts,
interdépendants les uns
des autres, ne
peuvent se
multiplier qu’avec de faibles chances de conserver leurs proportions
respectives initiales (pourtant nécessaires à une conservation dynamique). Dans
le « monde à ADN » qui lui a succédé, la cellule déleste en grande
partie les acides nucléiques de leurs activités catalytiques (assumées par des
protéines, non auto-réplicatives donc fortement dépendantes) mais en tire un
avantage considérable : L’ADN gagne en inertie (double hélice) et l’unidimensionnalité
propre aux acides nucléiques (combinaison linéaire de nucléotides) permet en se
prolongeant au delà des séquences utiles, d’unir sans
dommages une suc-
cession des
séquences utiles (gènes), de sorte que les propriétés auto-réplicatives de
l’ADN conserve les séquences mais aussi leurs proportions relatives.
Ainsi, du fait de ce fractionnement (fig.6), la non conservation
quantitative des stocks moléculaires finit obligatoirement (du fait de la
structure chromosomique de l’ADN) par rétablir un équilibre dynamique, par
exemple quand tous les stocks sont uniformément doublés. Doublement,
mitose ; saut qualitatif : Nous passons d’une cellule à deux cellules
conservant les caractéristiques structurales et fonctionnelles de la première.
Telles sont les bases de l’hérédité cellulaire.
Etapes du scénario de passage d’un monde à ARN à un monde à ADN : MP ; matière première, AR ;
auto-réplication, SP ; synthèse protéique, T ; transcription,
TI ; transcription inverse. Phase 1 : Les ARN présentent une double vertu ; celle
de s’auto-répliquer et celle de catalyser des réactions chimiques. Préalable théorique : rejet des notions de stéréo-spécificité
et de dualisme moléculaire.
Fig.6
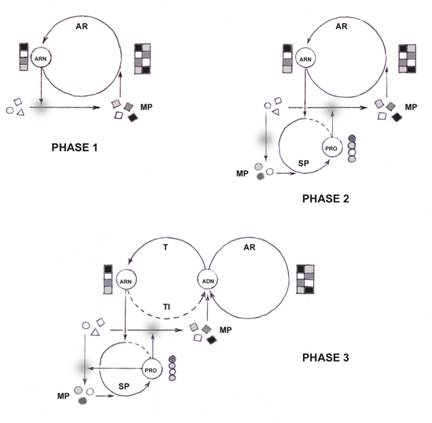
Concluons provisoirement. Les gènes sont bien des structures
auto-réplicatives, autonomes à l’origine, sur le modèle théorique de Richard
Dawkins : Les activités catalytiques qu’ils présentent n’ont d’autre
« fin » que de s’auto répliquer. Si les catalyses furent à l’origine
fortuites et totalement a-spécifiques avant la phase 1, c’est à dire
lorsqu’elles étaient assurées par des molécules inorganiques comme les argiles,
elles deviennent en fusionnant avec les molécules auto-réplicatives
elles-mêmes, l’objet d’une sélection naturelle d’autant plus puissante que la
multiplication des meilleurs réplicateurs explose. Cette sélection a pour effet
d’améliorer toujours d’avantage la stéréo-spécificité des réactions enzymatiques
jusqu’à la phase 3 ; A ce stade, la spécificité devient non pas
« maximale » comme le pensent les généticiens moléculaires, mais
« optimale » : Lorsque la « conservation » par
réplication supplante les effets de l’usure moléculaire, elle n’est plus une
simple compensation, mais un développement à la fois spatial et temporel.
b. Deuxième niveau : L’hérédité pluricellulaire
Parmi les types de divisions cellulaires connues actuellement, un détail
peut frapper ceux qui décrètent l’essence fondamentalement différente des uni
et des pluricellulaires : Nombreux sont les pluricellulaires primitifs
(« syncytium » comme les champignons basidiomycètes, de nombreuses
algues et animaux spongiaires) caractérisés par un développement cytoplasmique
et une multiplication des noyaux (caryocinèse) sans cloisonnements cellulaires
(cytodiérèses), de sorte qu’ils sont à la fois unicellulaires (une seule
cellule, volumineuse ou ramifiée) et pluricellulaires (nombreux noyaux).
Une fois encore, aucune logique n’associe a priori la cytodiérèse
(division de la cellule toute entière) à la caryocinèse (division du noyau
après duplication totale de l’ADN). Le développement cellulaire, inhérent comme
nous venons de le voir à la « conservation » cellulaire, est
matérialisé par la caryocinèse et conséquemment par le développement volumique
du cytoplasme. La cytodiérèse matérialise au départ la nécessité physique
d’assigner un noyau à chaque portion géographique du cytoplasme, de plus en
plus volumineux et dans lequel les noyaux doivent rester bien dispersés contre
la gravité ; l’interaction noyau – cytoplasme étant partout nécessaire à
l’activité cellulaire.
En conséquence l’association caryocinèse – cytodiérèse, c’est à dire la
mitose sous sa modalité la plus générale, est d’abord fortuite. Elle est
ensuite nécessaire moins à l’hérédité cellulaire (conservation dans le temps)
qu’à l’hérédité pluricellulaire, comme nous allons le voir.
Chez de nombreux insectes, mais aussi chez tous les oiseaux et tous les
reptiles, par exemple, la cellule-œuf commence ainsi par une série de
caryocinèses formant un syncytium, avant que l’embryon ne se
« cellularise ». Le développement embryonnaire, dans ses premières
étapes au moins et par suite d’un développement spatial inéluctable, peut être
assimilé à une conservation dans l’espace des propriétés moléculaires de la
cellule-œuf. Cependant le milieu dans lequel cette cellule-œuf se développe
n’est pas isotrope et son cytoplasme lui-même ne l’est pas : Dans la
mesure où les générations cellulaires provenant de cette cellule-œuf ne forment
pas une population (clone) d’unicellulaires isolés mais un ensemble de cellules
contraint par une « coquille » ou par une « zone
pellucide » à rester ensemble, l’embryon pluricellulaire n’aura d’autre alternative
que d’acquérir une structure hétérogène, adaptée en tout point au
micro-environnement dans lequel chaque cellule embryonnaire se tient
(non-conservation structurale). Détaillons les mécanismes qui sous-tendent
l’auto-dynamisme embryonnaire.
Première
contradiction antagoniste
Tout d’abord il est utile de rappeler que les modalités habituelles de
la reproduction sexuée ; gamétogenèse, méiose, fécondation, sont connues
chez la très grande majorité des unicellulaires eucaryotes actuels
(contrairement aux bactéries, les eucaryotes possèdent un noyau limité par une
enveloppe et des organites cytoplasmiques spécialisés), coexistant alors avec
d’autres modalités reproductives, asexuées. Chez ces espèces[10]
unicellulaires, les deux étapes de la reproduction sexuée (méiose et fécondation)
ne sont jamais associées dans un processus unique.
Un exemple parmi d’autres. Pluricellulaire primitif, Sordaria est
un champignon dont le cycle de développement est haploïde : L’organisme
(filaments de cellules non-spécialisées) est composé de cellules haploïdes
(chaque gène en un seul exemplaire). Lorsqu’il en rencontre un autre au cours
de sa progression (croissance), il forme des gamètes haploïdes. La rencontre de
deux gamètes forme une cellule « di-haploïde » (les deux noyaux ne fusionnent
pas en un seul). Cette dernière prolifère en un organisme di-haploïde, dont
certaines cellules seulement accompliront une caryogamie (fusion des
deux noyaux en un noyau diploïde ; fécondation tardive). De telles
cellules-œuf subissent enfin une méiose et forment des spores haploïdes. Une
fois libéré, chacun de ces spores germera en un nouvel organisme haploïde. Dans
ce cycle de développement, la reproduction sexuée existe, mais méiose et
caryogamie sont nettement dissociées dans le temps (comme pouvaient l’être
cytodiérèse et caryocinèse pour la mitose).
Qu’il s’agisse d’espèces haploïdes (méiose après caryogamie) ou
d’espèces diploïdes (caryogamie après méiose), ces deux étapes sont néanmoins
nécessairement liées l’une à l’autre pour conserver la ploïdie des organismes
au cours de leur cycle de développement. Celui des cellules bactériennes, les
plus primitives, indique clairement l’antériorité historique de la caryogamie
sur la méiose :
Chez les bactéries en effet, l’ADN ne forme qu’un long chromosome unique
(donc haploïde) mais il est très fréquent de détecter dans le cytoplasme de
petit ADN surnuméraires, facultatifs, appelés plasmides. Les plasmides
contiennent très peu de gènes et ne sont utiles à la cellule que lorsque le
milieu présente un danger ponctuel (gènes de résistance). Quand le danger se
présente, on assiste à des échanges tous azimut de plasmides entre cellules (et
ce même quand les bactéries n’appartiennent pas à la même espèce en
général) ; Le plasmide se duplique pendant que la cellule adhère à une
cellule voisine dépourvue du plasmide, la copie du plasmide lui est alors
transféré. L’ensemble du processus, très général chez les bactéries, est appelé
conjugaison bactérienne. Il offre à la colonie en péril des capacités de
résistance collective bien plus grande en milieu défavorable. Signalons que
cette résistance est le résultat d’un altruisme parfaitement hétérodoxe pour la
théorie néo-darwinienne de la sélection naturelle.
La conjugaison est le processus primitif annonciateur de la fécondation,
ce dernier consistant lui aussi à mettre en commun une plus grande
variété de gènes homologues
de façon à
augmenter les
chances de survie
cellulaire en milieu défavorable. Comme le phénotype découle autant de la
nature des gènes que de leur combinaison, leurs proportions relatives et leurs
interactions, la méiose qui suit cette fécondation atteste du fait qu’un simple
accroissement quantitatif du noyau, même par l’apport de nouvelles versions
géniques, ne suffit pas à rétablir un phénotype cohérent et conservé : La
cellule diploïde réduit le nombre de ses gènes et les recombine les uns par
rapport aux autres (saut qualitatif).
L’alternance méiose – fécondation, marquant la reproduction sexuée chez
toutes les espèces eucaryotes, uni comme pluricellulaires, haploïdes comme
diploïdes, semble donc naître primitivement d’une stratégie de conservation en
milieu défavorable, jouant sur les recombinaisons diversifiant les individus de
la population (méiose, responsable du polymorphisme et visant à augmenter les
chances de survie de certains individus au moins) autant que sur la mise en
commun des différentes variétés de gènes susceptibles de conférer à la
population une meilleure résistance au milieu (fécondation).
L’apparition de cette modalité semble avoir été complexe et fortuite.
L’exemple de la Paramécie, unicellulaire aquatique, illustre le caractère
apparemment fortuit des modalités primitives de la reproduction sexuée :
La Paramécie est « di-diploïde ». Elle possède un noyau actif
diploïde (noyau « somatique ») qui ne subit que des mitoses
(caryocinèses) et un noyau diploïde plus petit et non fusionné au premier
(noyau « germinal ») qui subit
(rarement et en conditions défavorables) la méiose, pour participer à des
échanges nucléaires complexes entre cellules. La fécondation est ici
primitivement un processus altruiste d’échange génétique et non un processus de
peuplement (augmentation démographique): Chez la Paramécie, la reproduction
sexuée ne fait que modifier mutuellement deux cellules, sans en créer de
nouvelles. Autre exemple tiré du règne végétal : Tandis que les herbacées
annuelles, soumises à des conditions très fluctuantes adoptent une stratégie de
reproduction sexuée fréquente sans investir dans un développement
de leur biomasse, les arbres quant à eux, soumis à un milieu stable (écosystème
forestier), adoptent une stratégie de biomasse (leur développement et
leur longévité peuvent être très grands) en investissant plus rarement dans la
reproduction sexuée. La reproduction sexuée est bien une stratégie
conservatrice de la population dans le temps, mais le développement individuel
(pluricellulaire) en est une autre[11].
Ajoutons enfin un dernier exemple montrant que les modalités de la
fécondation n’ont pas toujours été celles que nous reconnaissons chez la grande
majorité des eucaryotes actuels : De nombreux pluricellulaires primitifs,
dont les cellules ne sont pas ou peu différenciées les unes par rapport aux
autres (la conservation structurale de la cellule-œuf dans l’espace semble primitivement
presque réussir). Ainsi certaines éponges (animaux pluricellulaires à trois ou
quatre types de cellules spécialisées) la fécondation répond à des modalités
trophiques, préexistant chez les unicellulaires tels que les amibes, celles de
la phagocytose. L’ovocyte conserve des propriétés amiboïdes et phagocyte (capte
puis avale) le spermatozoïde d’un congénère. Là encore, des processus
préexistant pour d’autres fonctions, ici alimentaires donc indirectement liées
à l’auto-renouvellement, servent fortuitement une fonction nouvelle, celle de
la reproduction sexuée.
Résumons… Nous sommes partis du processus primitif de conjugaison
bactérienne, entrant dans le cadre de
la conservation temporelle
du groupe spécifique (le niveau
structural de l’organisme
pluri-
cellulaire est
shunté puisque les unicellulaires forment des groupes distincts, même si les
barrières d’espèces ne sont réellement affirmées que chez les unicellulaires
les plus évolués, à savoir les protistes eucaryotes), pour trouver son analogue
évolué dans la fécondation, volet primordial de la reproduction sexuée chez
tous les eucaryotes. La mise en commun des versions géniques de l’espèce
augmente les chances de survie de chacun en milieu défavorable.
Il reste que ce processus ne peut se suffire à lui même :
L’accroissement quantitatif de l’ADN d’une cellule ne peut lui être à terme que
préjudiciable, puisque l’existence de la cellule, de ses caractéristiques
structurales et fonctionnelles, repose sur l’état qualitatif du noyau (rapport
des gènes entre eux, etc.) autant sinon plus que sur son état quantitatif
(somme des gènes qui, pour les généticiens instructionnistes, détermine une
somme sans rapport entre eux de caractères phénotypiques). La méiose peut être
ainsi perçue comme une innovation de la matière vivante, au cours de l’histoire
de la vie, permettant initialement de rétablir l’haploïdie cellulaire (saut
qualitatif) après une fécondation (changement quantitatif).
La méiose est la solution d’une contradiction antagoniste : La
fécondation en tant que stratégie de conservation temporelle de la structure
cellulaire en milieu hostile, débouche sur une méiose, phénomène de
réduction de la
ploïdie mais aussi
de recombinaisons complexes
et diversifiantes,
modifiant le
génotype mais aussi le phénotype cellulaire[12].
La reproduction sexuée correspond à une stratégie de conservation des
propriétés cellulaires aboutissant à un changement de ces propriétés (nouvel
individu) !
Secondairement, chez les eucaryotes diploïdes[13]
(le cycle de développement est dominé par la diploïdie, seules les gamètes sont
provisoirement haploïdes) la reproduction sexuée se fonde sur l’inversion
apparente des processus de méiose (gamétogenèse) et de fécondation (formation
d’une cellule-œuf diploïde) sans changer ses modalités générales : Il
s’agit en fait non d’une inversion des deux processus mais d’un développement
temporel extrême de la phase diploïde (uniquement représentée par la
cellule-œuf chez les espèces haploïdes) au détriment de la phase haploïde (réduit
à la formation des gamètes chez les espèces diploïdes).
La conservation qualitative du type spécifique, chez les espèces
unicellulaires ou pluricellulaires, est incompatible avec des hybridations
interspécifiques : En ce sens les barrières d’espèce, loin d’assigner à
chacune une essence propre et immuable, sont plutôt des processus nécessaires à
l’hérédité du type (conservation du type), collatéraux à celui de la
reproduction sexuée : primitivement chez les bactéries sans reproduction
sexuée, les barrières d’espèces sont nous l’avons dit, peu étanches. Ainsi, et
ceci est particulièrement bien connu chez les pluricellulaires, ces barrières
correspondent à des luttes[14]
(opposées mais) analogues à la lutte contre l’autogamie, c’est à dire contre la
fécondation de deux gamètes provenant du même individu, contraire à la
vocation d’échange génétique de
la reproduction
sexuée. Trois
stratégies de lutte contre l’autogamie, sans filiations historiques apparentes
mais aboutissant au même résultat, sont aujourd’hui abondamment décrites et
illustrées :
-
Le dimorphisme sexuel résout de fait
le problème : Les individus étant morphologiquement et physiologiquement
mâle ou femelle, l’autofécondation est impossible. C’est la stratégie
majoritaire chez la grande majorité des animaux mais aussi chez de nombreuses
plantes « dioïques ». C’est un processus et non un état puisque la
sexualisation se construit au cours de l’ontogenèse : acquisition des
organes génitaux, du phénotype sexuel, de l’aptitude à se reproduire.
-
Lorsque les organismes sont
hermaphrodites, cas fréquent en particulier chez les végétaux, l’acquisition
des deux sexes est souvent dissociée dans le temps. L’organisme est alors, par
exemple, d’abord mâle puis femelle, de sorte que ses spermatozoïdes et ses ovules
ne puissent jamais coexister ni donc se rencontrer.
-
Quand l’hermaphrodite produit les
deux types de gamètes en même temps, cas fréquent lui aussi chez les végétaux
(plantes à fleurs hermaphrodites, à pistil et anthères), des mécanismes
d’incompatibilité plus raffinés permettent au pistil d’empêcher (c’est bien un
processus actif) le développement du pollen s’il provient du même organisme
parental (reconnaissance très proche sans doute, selon les botanistes, des
reconnaissances immunitaires chez les animaux). Ces mécanismes sont
analogues pour l’incompatibilité intra-spécifique (contre
l’autogamie) et l’incompatibilité
interspécifique (barrière d’espèce) : Ils sont donc diversifiant d’une
part ; contre l’autogamie, conservateurs d’autre part ; contre
l’hybridation interspécifique.
Objet de toute l’attention des généticiens instructionnistes puisque
c’est chez eux que l’embryogenèse est la mieux décrite en termes
instructionnistes, que les notions de lignées germinale et somatique sont les plus
utilisées, les animaux pratiquent la reproduction sexuée avec une grande
régularité, que le milieu soit favorable ou hostile. Ils illustrent ainsi le
détournement historique des armes d’une lutte pour la conservation typologique
collective et spatiale (primitivement au niveau unicellulaire) contre les
fluctuations du milieu vers une lutte individuelle pour la conservation
temporelle : un organisme pluricellulaire meurt mais laisse au moins un
descendant…
Voici donc comment se traduit cette première contradiction ; Chez
les unicellulaires primitifs la mitose est une lutte pour la conservation
temporelle et individuelle (renouvellement des générations cellulaires en tant
que populations) et la reproduction sexuée une lutte pour la conservation spatiale
et collective (homogénéisation des versions génétiques dans le groupe par une
mise en commun relative), alors que chez les pluricellulaires, la mitose est
une lutte pour la conservation spatiale individuelle (la cellule-œuf forme une
population « altruiste » conservant dans l’espace ses
caractéristiques cellulaires, sans le pouvoir absolument, pendant l’ontogenèse)
et la reproduction sexuée une lutte pour la conservation temporelle collective
(le polymorphisme permet de pérenniser le groupe spécifique dans le temps
malgré les fluctuations du milieu).
Deuxième contradiction antagoniste
Une contradiction relative au niveau cellulaire (propriété
auto-réplicative de l’ADN) jette les bases d’un niveau de développement plus
complexe, le niveau pluricellulaire, dépassement du premier sur le plan
quantitatif. Suit une seconde contradiction sur le plan qualitatif, couvrant à
la fois deux mouvements auto-dynamiques ; Celui de l’auto dynamisme
ontogénétique (embryogenèse et développement de l’individu juvénile puis adulte),
et celui de la reproduction de cette ontogenèse d’une génération à l’autre
(hérédité pluricellulaire).
Considérons d’abord ce dernier mouvement. Ce qui était la solution
conservatrice d’une modification quantitative du matériel génétique cellulaire
(caryogamie) devient chez les pluricellulaires diploïdes, c’est à dire quand
elle « précède » cette caryogamie, le moteur d’un bouleversement
cellulaire : La méiose modifie la nature de cellules qui appartenaient
initialement à l’organisme parental (gamétogenèse). Elle est en ce sens la
ligne de démarcation entre le somatique et le germinal. Aucune
barrière physique entre « lignée » somatique éphémère et
« lignée » germinale immortelle, comme Weismann le postulait en son
temps. C’est un processus qui produit à partir des cellules d’un individu
(somatiques) de nouvelles cellules (germinales), les gamètes, qui ne lui
appartiennent plus.
Il n’est pas facile de désigner en quoi une cellule appartient ou non à
un organisme pluricellulaire. Une certitude ; Cela tient à sa structure
générale, plutôt qu’à ses seuls gènes. Conventionnellement une cellule ou un
tissu appartient au « soi » quand il n’est pas détruit par le système
immunitaire de l’organisme[15].
Nous conviendrons donc que les globules rouges (cytoplasme sans ADN), les
globules blancs (génétiquement différents entre eux dans le sang d’un même
individu) appartiennent à l’organisme qui les contient, qu’en revanche les
gamètes, résultant d’une méiose, ne lui appartiennent plus.
Deux remarques. Tout d’abord, dans la mesure où la distinction somatique
/ germinal peut être dialectique (elle entre dans le champs de notre définition
dialectique de la matière vivante), le concept
de lignée ne l’est en aucun cas. Tout biologiste en effet qualifie les
cellules souches qui s’apprêtent à subir la méiose, de
« germinales », comme les gamètes. Il a tort. Ces cellules souches
appartiennent encore objectivement au soi et sont donc somatiques tant qu’elles
n’ont pas subi la première division de méiose. Pas de lignée, développée continûment
de cycle en cycle indépendamment d’une lignée somatique lui restant soumise,
mais une répétition de sauts qualitatifs (méioses produisant le germinal),
répétition qu’on inférera à celle des rencontres entre gamètes (à un autre
niveau, celui de la première contradiction antagoniste). Une nouveauté
donc : Ce n’est pas la fécondation qui crée un nouvel
« individu » mais la méiose, antérieurement ; L’événement
contingent (rencontre des gamètes) est rapporté à un événement auto-dynamique
parental nécessaire.
Deuxième remarque. Cet événement autodynamique parental, qui n’est autre
que la gamétogenèse[16],
dépend à son tour de l’événement « extérieur » à l’organisme qu’est
la fécondation des gamètes. Dans le cas général d’une rencontre entre ovule et
spermatozoïde, notons que si le spermatozoïde ne devient pas seul un embryon
pluricellulaire, c’est qu’avec son noyau, il ne contient quasiment pas de
cytoplasme (or l’interaction noyau-cytoplasme est comme nous le verrons
indispensable à cette évolution), si l’ovocyte ne subit pas d’embryogenèse
avant sa rencontre avec un spermatozoïde, c’est qu’il est « bloqué »
(par des interactions avec le reste de l’organisme maternel) en cours de méiose
et que cette méiose ne se termine qu’au dernier moment : c’est le contact
avec un spermatozoïde qui re-déclenche et achève la méiose juste avant la
caryogamie. Cas d’école fort instructif : Toutes les ouvrières d’une ruche
naissent du développement spontané d’ovules non fécondés pondus par la
reine !
Pas de blocage en cours de gamétogenèse en revanche chez les diploïdes
unicellulaires et pluricellulaires primitifs : Dans ce cas, sans rencontre
entre deux organismes distincts, pas de gamétogenèse. La fécondation chez les
pluricellulaires évolués, suivant les mêmes
modalités à un détail
prés (la gamétogenèse finit par
anticiper la rencontre dans le cycle de développement), n’est donc contingente
qu’en apparence. Lorsque l’ovocyte est bloqué en méiose en attendant la
rencontre avec un spermatozoïde, nous dirons qu’il n’est pas une cellule
« vivante » proprement dite (en tant qu’il n’évolue plus qu’au
ralenti) mais une structure « léthargique » maintenue en vie du seul
fait d’interventions extérieures, interventions de l’organisme pluricellulaire
maternel. Blocage apparemment anti-dialectique, mais objectivement lutte active
d’un organisme pluricellulaire contre le développement d’une structure
étrangère en son sein.
Venons en au mouvement auto-dynamique le plus problématique en biologie,
celui de l’ontogenèse pluricellulaire. Voici précisément la jonction entre deux
niveaux, le niveau unicellulaire (gamètes, cellule-œuf) et le niveau
pluricellulaire (organismes parentaux, embryon). Plusieurs observations seront
utiles à l’élucidation de ce problème sous l’angle dialectique.
Balayons d’abord les sous-entendus idéalistes concernant une prétendue
prédestination génétique de la cellule-œuf à construire un organisme déterminé
(phénotype). Chez les végétaux, la morphologie de l’organisme est en perpétuel
changement au cours de sa vie et dérive de contraintes environnementales
évidentes d’une part (tropismes), de corrélations morphogénétiques d’autre
part (le
feuillage se
développe en fonction du développement racinaire et inversement). Chez les
animaux, qui apparemment répondent à des lois différentes (prédétermination
apparente des différentes parties du corps), l’ontogenèse passe par une série
d’états structuralement très différents (qui chez l’Homme, du fait de sa
viviparité, passent sous silence, donnant à
penser que le fœtus n’est
qu’un état latent
et sans individualité). Les
exemples en sont nombreux. La grenouille, insectivore et aérienne, provient de
la métamorphose d’un têtard végétarien et aquatique. Le papillon, volant et
consommateur de nectar, provient de la métamorphose d’une chenille, rampant et
herbivore. Mieux encore, l’ascidie, sorte de « sac » végétatif fixé à
un substrat, dépourvu de système nerveux, provient de la métamorphose d’une
sorte de têtard très mobile, doté d’une tête, d’une queue, de muscles et de
nerfs. La sacculine, parasite du crabe composé d’une simple poche pleine de
gamètes et d’un fin chevelu filamenteux en prolifération dans les organes de
son hôte, est en fait un crustacé lui même provenant de la métamorphose d’une
larve nageuse complexe et active, de type crevette. Quand une cellule-œuf forme
un embryon qui forme lui-même un têtard se métamorphosant finalement en
grenouille, le qualificatif d’être vivant individualisé convient sans conteste
à la grenouille autant qu’au têtard. Pourquoi en priver l’embryon et la
cellule-œuf elle-même ? Prétendre comme le font Dawkins ou Sonigo, que nos
gènes ou nos cellules vivent à travers nous, revient à affirmer que le têtard
continue de vivre à l’intérieur de la grenouille qu’il est devenu, que la chenille
se cache encore à l’intérieur du papillon !
Ainsi, non seulement la structure cellulaire de l’œuf ne peut se
conserver dans l’espace (différenciation des cellules au cours de
l’embryogenèse par exemple) mais encore la structure pluricellulaire, résultat
de cet échec, ne le peut pas davantage : témoins les métamorphoses, mues,
phénomènes chrono-biologiques divers chez les animaux, croissance et
développement permanent chez les végétaux…
En conséquence, la cellule-œuf ne recèle pas le programme permettant de
construire un organisme fini, c’est une structure qui se suffit à elle-même,
mais qui représente en tant que matière vivante, la base matérielle d’un
processus auto-dynamique de formes pluricellulaires successives qui la nient
puis se nient elles même. Si chaque forme se conserve d’une certaine façon dans
le temps, réapparaissant à un moment précis de l’ontogenèse à chaque
génération, c’est que l’ensemble du processus ontogénétique se conserve, la
voie par laquelle un ensemble de propriétés intracellulaires se conserve sur le
mode pluricellulaire. La formule d’Engels sur la réciprocité des causes et des
effets à un niveau supérieur à celui de l’analyse réductionniste (cf.
conclusion du chapitre précédent) prend ici tout son sens. Pourquoi la cellule-œuf,
structure unicellulaire, forme t-elle par mitoses un organisme pluricellulaire
dont les cellules inter-agissent étroitement et nécessairement, plutôt qu’une
population homogène de cellules-œufs isolées ? Parce qu’elle est
prisonnière pendant un temps suffisant d’une « coque » structurée
forçant les cellules à rester ensemble. Pourquoi la cellule-œuf est-elle
prisonnière d’une coque structurée ? Parce qu’elle résulte d’une
construction exogène (l’organisme maternel est l’environnement de l’ovocyte qui
ne lui appartient plus) organisée par les interactions existant entre les
cellules somatiques des organismes parentaux.
Si la viviparité, processus par lequel la cellule-œuf subit une
assistance prolongée pendant son développement, est un mode particulièrement
évolué chez les pluricellulaires, l’oviparité en reste le mode le plus
représentatif. Dans ce dernier cas la cellule-œuf est directement livrée au
milieu et y subit son développement sans assistance biologique apparente. Même
ainsi cependant, sa structure peut livrer quelques uns des mystères de son
auto-dynamisme. Elle correspond toujours à la confrontation intracellulaire
entre un noyau néoformé, possédant des caractéristiques nouvelles et un
cytoplasme préexistant, complexe, asymétrique et exclusivement construit par
l’organisme maternel (dont le matériel génétique n’est pas celui de la
cellule-œuf). Manifestement, une contradiction interne anime cette cellule très
particulière ; une sorte d’inadéquation originelle entre noyau et
cytoplasme. Le matériel enzymatique ovocytaire (maternel) permet les
duplications du noyau. Ce faisant, ne s’agissant pas de noyaux maternels, ces
duplications engendrent une modification cytoplasmique pendant les
renouvellements moléculaires. L’hérédité pluricellulaire, c’est à dire la
reproduction du processus ontogénique, reposerait alors partiellement sur
l’interaction contradictoire initiale noyau/cytoplasme et non sur des
« gènes architectes » du noyau seul (ces gènes existent mais n’ont
pas pour « vocation » de programmer le développement
embryonnaire) : La théorie préformiste doit être rejetée (qu’il s’agisse
du modèle fantaisiste des « poupées russes » au XIXe
siècle ou des gènes architectes aujourd’hui), au profit d’une théorie
épigénétique (succession d’étapes dont chacune est induite par la précédente)
conforme à la dialectique matérialiste.
Développons ce point capital. La biologie du développement est
aujourd’hui sujette à une contradiction bien étrange. Pratiquement, l’épigenèse
s’est totalement substituée (à la lumière des résultats expérimentaux) à la
préformation, en introduisant un terme nouveau, celui de totipotence, que nous
développerons. Théoriquement, la génétique instructionniste prévaut toujours,
expliquant les asymétries embryonnaires (différenciations cellulaires) par des
interactions signalétiques (les « signaux » chimiques et les
récepteurs moléculaires spécifiques de ces signaux étant déterminés par des
gènes) : Le futur dos reçoit un signal du futur ventre lui imposant de se
« dorsaliser » et réciproquement. Ainsi une asymétrie présomptive
(présence de récepteurs dans les futures cellules dorsales, de signaux dans les
futures cellules ventrales) préexiste (préformation) à la réalisation de
l’asymétrie.
Sortons de cette contradiction apparente. Tout d’abord, la cellule-œuf
n’est jamais homogène, comme cela a été dit. Elle hérite d’une structure
ovocytaire complexe façonnée par l’organisme maternel (pellicule solide,
gangue, couronne de cellules nourricières, noyau excentré, anisotropie
cytoplasmique du vitellus –réserves-). D’une certaine façon, elle est donc
effectivement préformée. Cependant cette forme ne correspond en rien à celle de
l’organisme final. Contre la théorie épigénétique, nous dirons que le
développement embryonnaire tient au moins autant à la structure ovocytaire
façonnée par l’organisme pluricellulaire maternel (« effet
maternel », dans un premier temps) qu’aux potentialités du noyau de la
cellule-œuf (« effet zygotique », dans un second temps). Contre la
théorie préformiste, nous dirons que l’embryogenèse est un processus déterminé
d’inductions successives (épigenèse) résultant, sans programme préexistant,
d’une contradiction entre la tendance des noyaux à réaliser des structures
cellulaires propres et l’action hétérogène et contraignant du milieu
extracellulaire (extra et intraembryonnaire).
Aucun embryologiste ne peut plus nier le rôle moteur des asymétries de
la cellule-œuf sur son devenir. Citons le rôle nécessaire et suffisant des
croissants pigmentaires (batraciens, échinodermes), de la gravité et de la
forme de l’œuf (insectes, oiseaux), de la disposition cytoplasmique du
vitellus, du noyau (tous les animaux), de la disposition hétérogène et orientée
des cellules nourricières, folliculaires autour de l’ovocyte, dans
l’acquisition embryonnaire ultérieure des axes de polarité antéro-postérieur et
dorso-ventral. La centrifugation d’une cellule-œuf, provoquant
l’homogénéisation de son cytoplasme, fait de lui une forme embryonnaire
monstrueuse cessant très précocement son développement.
D’une façon extrêmement générale chez les animaux, la cellule-œuf forme
tout d’abord un ensemble sphérique de cellules plus ou moins semblables, de
plus en plus nombreuses et (puisque l’embryon, dans sa coque, conserve le même
volume) de plus en plus petites. Sans sa coquille d’ailleurs, aucun œuf ne peut
se développer. L’asymétrie cytoplasmique, généralement due à un gradient
vitellin (plus le cytoplasme est dense en vitellus, plus
sa segmentation mitotique
sera difficile : C’est particulièrement évident pour
l’ « énorme » cellule-œuf d’oiseau –ou jaune-
très riche en
vitellus, qui ne se
segmente qu’en périphérie,
à l’endroit qu’occupait
initialement le noyau zygotique)[17],
explique que cette « morula » présente des petites cellules d’un côté
(peu de vitellus ; pôle animal) de grosses cellules de l’autre (beaucoup
de vitellus ; pôle végétatif). Cette variété de taille est à l’origine
d’une cavitation centrale spontanée (étape « blastula »).
L’embryon continue sa segmentation mais ses cellules qui atteignent une
taille minimale limite, sont forcées de former localement une invagination, un
pli vers la cavité centrale, encore vide de cellule (la coque existe toujours à
l’extérieur). Ce phénomène, formant une « gastrula » correspond à la
première étape d’une mise en place des feuillets embryonnaires (début de
différenciation) : Les cellules restant à l’extérieur formeront
l’ectoderme (future peau, système nerveux, etc.), les cellules invaginées par
l’embolie formeront l’endoderme (tube digestif, glandes digestives) et le
mésoderme. Les embryogenèses se diversifient d’espèce en espèce, chacune
poursuit les étapes en allant comme on le voit, du simple au complexe. Les
premières étapes du développement décrites ici sont elles mêmes dans la forme,
très différentes d’une espèce à l’autre.
Ainsi le rôle des contraintes physiques s’opposant au développement
spontané de l’embryon pluricellulaire apparaît au moins aussi important que des
mécanismes chimiques qui guideraient l’activité des cellules et émanant d’une
expression génétique. Asymétries et contraintes physiques initiales sont
primordiaux, en tant qu’antagonistes au développement cellulaire spontané.
L’extrême complexité structurale et fonctionnelle de l’embryon puis de l’adulte
ne résulte pas d’une extrême complexité préexistante au niveau des gènes, mais
de la complexification d’une structure « simple »[18],
la cellule-œuf.
Pourquoi sommes nous toujours tentés de considérer la forme adulte comme
la forme finie, prévue d’avance par un programme, et les formes embryonnaires
comme de simples étapes entre
deux
générations
successives d’organismes adultes ? Parce que les étapes embryonnaires sont
très rapides et que la forme adulte est bien plus longue (mieux conservée dans
le temps). Les embryologistes affirment : Dans les premières étapes de
l’embryogenèse, l’expression de certains gènes (bien sûr non encore
identifiés !) impose aux cellules de se diviser très vite. Lorsque les
organes sont différenciés en cellules hépatiques, cardiaques, intestinales, etc.,
leur cycle de division est fortement ralenti et leurs mitoses, à cause de
répressions génétique par différents facteurs de transcription, sont beaucoup
plus rares. Retournons l’idée ! Les cellules embryonnaires, dans un
micro-environnement plein de contraintes physico-chimiques, ne peuvent
conserver leur structure longtemps : Elles se divisent donc très vite, la
mitose étant la solution d’une telle contradiction (voir chapitre précédent).
Par suite de la complexification, les interactions « altruistes » des
cellules, nécessitant des asymétries cellulaires inévitables ou différenciations,
finissent par se substituer en tant que « milieu »
extracellulaire-intraembryonnaire au milieu extraembryonnaire plus simple
d’origine. Ce nouveau milieu, très propice à la conservation de cette nouvelle
forme pluricellulaire, s’oppose beaucoup moins à la conservation particulière
de chaque cellule : Celles ci se divisent donc beaucoup moins (vie plus
longue).
On passe progressivement de quelques cellules « totipotentes »
(c’est à dire capables de devenir n’importe
quel type de
cellules différenciées si on les délocalise artificiellement sur
l’embryon) à de très
nombreuses
cellules différenciées (stade de conservation globale de la forme
pluricellulaire, résultant de l’échec de la conservation zygotique) : Là
encore, c’est un saut qualitatif évident.
Parmi les potentialités cellulaires, qui préexistaient historiquement au
stade pluricellulaire, celle d’accomplir
la méiose peut
être laissée à
certaines cellules de
l’organisme. On appellera celles-ci « cellules souches des
gamètes ». Elles finissent par être, génétiquement au moins, distinctes
des cellules somatiques au sein desquelles elles
vivent, s’en trouvent isolées
par une coque et s’enrichissent
en réserves pour survivre. C’est donc encore fortuitement, c’est à dire sans
« préméditation », que se réunissent les conditions permettant le
développement embryonnaire de la cellule-œuf[19].
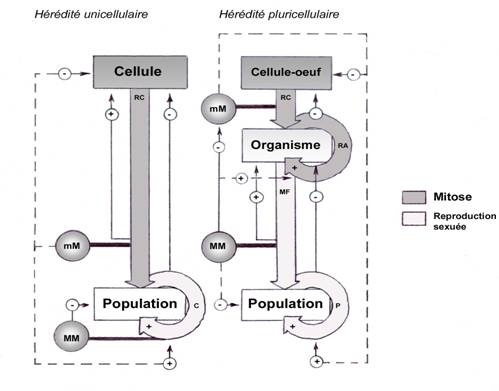
Fig.7 ;
Processus héréditaires chez un unicellulaire et chez un pluricellulaire.
mM ; micromutations (mutations ponctuelles changeant le nombre et l’ordre
des nucléotides dans un gène), MM macromutations (mutations chromosomiques
changeant le nombre et l’ordre des gènes dans le génome), RC ;
reproduction conforme, RA ; régénérations-différenciations-accomodations,
MF ; méiose et fécondation (conservation du caryotype), C ;
conjugaisons, P ; polymorphisme.
D’une façon générale, les processus conservateurs cellulaires, autrement
dit l’hérédité cellulaire, correspondent à des innovations dépendantes
des micromutations s’accumulant dans les gènes, mais se chargent de lutter
activement contre ces micromutations. Les processus conservateurs
pluricellulaires, autrement dit l’hérédité pluricellulaire,
correspondent à des innovations dépendantes à la fois des micromutations et des
macromutations ; ces processus intègrent la macromutabilité comme base de
l’hérédité (ontogenèse) tout en continuant de lutter contre la micromutabilité.
Moteur du polymorphisme au niveau du groupe spécifique, cette dernière reste
toutefois utile et ne peut disparaître totalement (fig.7)
Notre phénotype humain ne résulte donc pas d’une destruction
tendancielle des processus conservateurs du niveau cellulaire, comme cela était
proposé en conclusion du chapitre précédent, mais plutôt d’une négation de ces
processus ; Notre organisme reflète la négation héréditaire des activités
de nos gènes, par l’activité de processus d’ordre supérieur. Etant héréditaire,
reproductible, cette négation ne détruit pas les gènes, elle les conserve
obligatoirement et autant que possible : l’appareil reproducteur est le
lieu où cette conservation de l’hérédité cellulaire s’avère la plus propice.
B. Bilan et critique destinée à une amélioration théorique
Le matériau de notre synthèse est théorique, autant que pratique :
La science accusée, qui a nom « Théorie synthétique de l’Evolution »,
fournit 1) des résultats expérimentaux très variés et très documentés, même
s’ils sont « orientés » et souvent travestis par des présuppositions
idéologiques, 2) un système conceptuel riche, fruit d’au moins quarante ans de
génétique moléculaire, ce qui représente effectivement peu, science qu’on
n’hésitera pas à qualifier de bourgeoise, eu égard à son caractère idéaliste,
métaphysique et à l’investissement financier, politique et médiatique dont elle
bénéficie. Notre problématique est la suivante ; Il s’agit de réévaluer ce
système conceptuel à la lumière d’une méthode qui, en d’autres domaines a fait
ses preuves, le matérialisme dialectique.
Produit-on des vérités objectives par un travail théorique de ce
type ? La question est mal posée. Les vérités objectives partent de la
pratique, c’est à dire en première instance des résultats expérimentaux. Si
l’existence d’une séparation étanche entre les cellules germinales et les
cellules somatiques était prouvée par la pratique, nous l’admettrions. Or ce
n’est pas le cas. La connaissance des théories actuelles montre qu’elles ne
sortent pas « positivement » des résultats, mais qu’elles cherchent plutôt
stratégiquement à s’en accommoder[20].
L’histoire des sciences témoigne que cette accommodation a
d’abord été infructueuse[21],
elle a connu ensuite une assez longue période de succès et d’enthousiasme
théorique, qui s’achève aujourd’hui.
Pour autant, doit-on tout rejeter de cette science bourgeoise, à
l’instar des savants occidentaux qui ont tout rejeté du lyssenkisme ?
Certes non ! Marx et Engels ont-ils tout rejeté de Hegel ? Ont-ils
tout rejeté de Darwin, après avoir désigné précisément ce qui en faisait un
savant bourgeois ? En tant que matérialiste, on se doit d’évaluer en toute
conscience des tenants et aboutissants métaphysiques de la génétique
moléculaire, le bien fondé des instruments de cette théorie, avant d’en
effectuer éventuellement le renversement.
« Analyser
la nature en ses parties aliquotes, répartir les divers phénomènes et objets
naturels en classes déterminées, scruter la constitution interne des corps
organiques selon leurs multiples formes anatomiques, c’étaient là les
conditions essentielles des progrès gigantesques que nous ont apportés ces
quatre derniers siècles dans la connaissance de la nature. Seulement tout cela
nous a aussi laissé l’habitude de considérer les objets et les phénomènes de la
nature isolément, en dehors de leur grande connexion d’ensemble ; et, par
conséquent, non pas dans leur mouvement, mais à l’état de repos, non pas comme
essentiellement changeants, mais comme fixes, non pas dans leur vie, mais dans
leur mort. Et cette conception, transportée, comme elle l’a été par Bacon et
Locke, de la science de la nature dans la philosophie, y créa l’étroitesse
scientifique des derniers siècles, le mode de pensée métaphysique. »
F.Engels. Anti-Dühring.
Reste à traiter avec précaution ce qui peut ici apparaître comme une
forme de relativisme. Nous n’avançons pas masqués : Le parti pris,
conforme aux lois du matérialisme dialectique et entretenant un certain rapport
à une pratique qui est réelle certes, mais qui n’est pas la nôtre, c’est celui
d’éviter les polémiques théoriques au cours de l’analyse pour mieux en
retrouver le poids au terme de celle-ci.
A conserver a priori : La ségrégation mendélienne des effets
phénotypiques, pas le mendélisme (à cet égard, Timiriazev et Mitchourine en
firent autant), les notions de germen, de soma, pas le weismanisme (notion
idéaliste de lignées sans interaction), la réalité des succès de Mitchourine et
de Lyssenko, pas le Lyssenkisme (sa critique du mendélo-weismanisme mise à
part), les notions de gène, d’ARN, etc., pas la génétique moléculaire (dogme
central). L’important étant de considérer ces formes, qui sont bien des formes
matérielles, dans leur évolution et non dans leur structure apparemment fixe,
en prenant garde qu’un nouvel agnosticisme ne surgisse de ce relativisme.
« Toutes les
anciennes vérités de la physique, y compris celles qui furent considérées comme
immuables et non sujettes à caution, se sont révélées relatives ; C’est donc qu’il ne peut y avoir aucune vérité objective indépendante
de l’Humanité. Telle est l’idée non seulement de toute la doctrine de Mach mais
aussi de tout l’idéalisme « physique » en général. Que la vérité
absolue résulte de la somme des vérités
relatives en voie de développement ;
que ces reflets
deviennent de plus en plus exacts ; que chaque vérité scientifique
contient en dépit de sa relativité un élément de vérité absolue, -toutes ces
propositions évidentes pour quiconque a réfléchi à l’Anti-Dühring d’Engels, sont de l’hébreu pour la théorie
« contemporaine » de la connaissance. »
Lénine. Matérialisme et Empiriocriticisme.
Les gènes existent, mais la génétique mendélienne agonise… Voilà qui
surprendrait sans doute un Schmalhausen … un Lyssenko à plus forte raison. Le
moment est venu de regarder en face la matière vivante, aussi mal décrite
fut-elle, et de montrer aux gardiens du dogme ce qu’ils ignorent
eux-même : Leurs reculs incessants, qu’ils vivent comme de simples
compromis sans enjeu, et la compatibilité de leurs instruments théoriques
fondamentaux avec le matérialisme dialectique. Une fois la ligne de démarcation
tracée entre ce qu’ils ont objectivement découvert et l’exploitation idéaliste
qu’ils en ont faite, cette dernière n’en aura jamais été aussi
vulnérable !
« La science
prolétarienne ne peut, sans se mutiler, rejeter tout le contenu de la science
bourgeoise, mais doit tirer, dans ce contenu, ce qui doit enrichir sa propre
théorie »
M.Prenant. Biologie et Marxisme 1935
Mais cette ligne n’est ici qu’esquissée… partant d’une hypothèse de
travail, notre définition de la matière vivante, qu’il s’agira de
perfectionner. N’oublions pas que si des lois dialectiques émergent
maladroitement et partiellement des théories idéalistes actuelles, c’est
qu’elles constituent les « grains » d’objectivité de ces théories
existant en fait dans la matière avant de s’élever au niveau théorique.
De même les distinctions soma-germen, gènes-protéines, les barrières d’espèces,
sont des concepts idéalistes mais qui émergent d’une réalité concrète de la
matière comme des processus conservés à travers l’histoire de la vie :
Notre définition de la matière vivante n’explique pas seulement ce qui
caractérise la vie, elle explique aussi pourquoi il est (était) si facile de
donner de cette matière des comptes rendus idéalistes lorsqu’elle est observée
empiriquement, sans théorie matérialiste préalable… Parallèlement à son utilité
purement scientifique, notre définition est donc une vigoureuse opposition à
l’empirisme et au formalisme autant qu’un remède contre l’idéalisme surgissant
de tout matérialisme non-dialectique.
La théorie de Darwin indique que le moteur de l’Evolution est le couple
hasard – sélection. Sans oublier l’étroite relation qui existe entre le hasard
(mutations « aveugles ») et la sélection (sélection naturelle
« aveugle ») et le caractère provisoire de la formulation
darwinienne, nous reprendrons ces concepts successivement, par soucis de
clarté.
a. Le statut du hasard et la question de l’autogenèse
Tout en étant conforme à la dialectique, le principe de causalité, relié
à celui de nécessité, doit être manié avec précaution.
« Le
caractère universel et multiple des interactions dans le monde, la causalité ne
l’exprime que de façon unilatérale, partielle et incomplète ».
Lénine. Matérialisme et Empiriocriticisme
La conscience des lois de la pensée dialectique permet de renvoyer dos à
dos le finalisme, qu’il s’agisse d’un finalisme réductionniste ou
spiritualiste, et l’indéterminisme ; définition agnostique voire
franchement spiritualiste du hasard. La littérature scientifique propose
différentes définitions du hasard des plus idéalistes aux plus
matérialistes ; Causalité « ultra-complexe », rencontre de
séries causales indépendantes, contingence (événement non nécessaire),
surdétermination, hasard statistique, chaos déterministe. Admettons que la
matière ne saurait être animée dans son mouvement d’une nécessité universelle,
puisqu’elle n’est auto-dynamique que dans ses parties interactives, pas dans la
totalité, qui recoupe ces parties sans discrimination qualitative. Ainsi le
mode de production socialiste se substituera au mode de production capitaliste
(mouvement autodynamique), à moins qu’un météorite géant n’extermine l’Humanité
entre temps ! La trajectoire du météorite rend l’extermination de
l’Humanité nécessaire, le mode de production capitaliste rend également
nécessaire son passage au socialisme. On ne peut parler pour autant de
nécessité du capitalisme à périr d’une telle catastrophe ! …
« Pas plus
que la quantité et la qualité, le hasard et la causalité ne s’opposent en
antithèses rigides »
Prenant.Biologie et Marxisme
« Le hasard
n’est que l’un des pôles d’un ensemble dont l’autre pôle s’appelle
nécessité »
F.Engels.
Origine de la famille, de la propriété privée et de l’état
C’est en ce sens que nous interprétons les transformations
« fortuites », collatérales à des transformations
auto-dynamiques : passage d’un niveau structural à un autre, innovations
structurales, diversité des formes coexistantes de la matière vivante.
Mais c’est au niveau moléculaire, niveau invisible pour l’œil humain,
donc pour lequel les tentations métaphysiques sont légion, que la notion de
hasard est particulièrement polémique ; Les variations structurales
individuelles naissent-elles de mutations génétiques neutres au hasard ?
Le problème est celui de l’indéterminisme. La génétique moléculaire affirme que
les mutations apparaissent régulièrement, accidentellement, indépendamment du
milieu et de façon non-orientée. Non, les mutations n’apparaissent pas
indépendamment du milieu. Mais ; oui, elles apparaissent de façon
non-orientée…
Evolution génétique (accumulation d’erreurs pendant les duplications
successives) et action du milieu ne peuvent en aucun cas être
indépendantes ; Matières vivante et non vivante sont en interaction
permanente et complexe. Ainsi, lors d’un stress environnemental, les bactéries
accélèrent prodigieusement le rythme de leurs mutations génétiques en particulier
sur les gènes déterminant les protéines rendues inutiles dans ce nouveau
contexte : Les mécanismes conservateurs de la matière vivante sont donc antérieurs
aux mutations, ils en sont l’une des conditions matérielles. La rapidité
d’acquisition d’une résistance à un insecticide par une population de moustique
(environ un an), montre que ces mécanismes actifs d’adaptation rapide ne
concernent pas que les unicellulaires, dont la multiplication est suffisamment
rapide pour surseoir à la lenteur supposée des mutations génétiques spontanées.
Les exemples d’adaptation pluricellulaire sont nombreux, notamment en
parasitologie. Ajoutons que ces mécanismes sont étroitement associés aux
conditions environnementales, qui sont corrélativement les conditions de
fonctionnement de la cybernétique cellulaire.
Dans l’exemple d’une colonie bactérienne sensible soumise à un
antibiotique, les néodarwiniens affirment, pour expliquer l’adaptation, que la
forme résistante est déjà représentée, et ce de façon ultraminoritaire, avant
l’exposition à l’antibiotique, devient majoritaire (déplacement du phénotype
moyen de la population vers cette forme, après plusieurs générations
cellulaires) par la sélection naturelle. Les néo-lamarckiens affirment quant à
eux que cette résistance ne préexiste pas et que le phénotype moyen s’y déplace
de façon orientée lorsque la colonie est soumise à l’antibiotique. La situation
réelle est la suivante : La résistance ne préexiste effectivement pas.
Lorsque la colonie est soumise à l’antibiotique, le polymorphisme s’élargit
fortement, mais de façon non-orientée, autour du phénotype moyen
sensible ; L’une des formes créées correspond alors à la résistance.
L’évolution, seulement alors, suit le modèle darwinien précédemment cité [voir
schéma B.c].
Concernant les propriétés conservatrices de la cellule, rappelons deux
éléments fondamentaux :
1. C’est la propriété auto-réplicative de l’ADN, inférée par sa
structure chimique, qui se tient à la base des propriétés conservatrices plus
ou moins raffinées de la matière vivante, dans la mesure où les gènes
contrôlent le renouvellement des molécules actives de la cellule. Les mutations
génétiques ont mécaniquement des effets multiples, avantageux ou délétères, sur
la cybernétique cellulaire. Le contrôle des gènes sur les synthèses protéiques
implique une conservation du code génétique. Celui-ci apparaît fortement
anti-dialectique, puisqu’il est universel, immuable et arbitraire. Ce n’est pas
le cas [voir annexe « Code génétique et spiritualisme »].
2. Si les gènes dominent la synthèse protéique, il faut admettre, à la
lumière du cas considéré plus haut, que le milieu (pour les gènes, il s’agit,
ne l’oublions pas, du milieu extracellulaire –l’antibiotique, qui passe
d’ailleurs dans le cytoplasme- et du milieu intracellulaire non nucléique
–protéines-) peut à son tour les dominer. Ceci n’est concevable que dans la
mesure où il existe une cybernétique cellulaire, émergeant de la réciprocité
des interactions entre toutes les molécules cellulaires. Envisageons cette
cybernétique comme un ensemble complexe de contraintes fonctionnelles, garant
d’un niveau de conservation supérieur à celui des gènes (a priori la moindre
variation remet en cause la totalité de cette cybernétique cellulaire, cette
dernière se conserve globalement ou s’avère létale sans condition). Il
existerait une « loi de corrélation des molécules » apparentée à la
« loi de corrélation des organes ».
Il est d’ores et déjà évident que la distinction problématique
autogenèse – ectogenèse en ce qui concerne l’évolution de la matière vivante
est illusoire et métaphysique. En revanche, un problème résiste encore à
l’analyse : La matière vivante est-elle le siège d’une conservation de structure,
comme l’indique notre hypothèse de travail, ou d’une conservation de processus ?
Il est facile de localiser ce problème ; Il se tient dans la définition
dialectique de la matière. On peut avoir l’impression par exemple que
l’hérédité cellulaire consiste en une conservation structurale tandis que
l’hérédité pluricellulaire consiste en une conservation d’un processus
ontogénique. Mais, de même que la matière ne saurait être inerte, le mouvement
(processus) ne saurait être envisagé sans matière. Structure, processus, nous
parlons bien de la même chose. C’est le lien nécessaire entre ces deux
hérédités qualitativement distinctes qui reste problématique. Signalons
qu’elles sont différées dans le temps, et que leur causalité ne peut être
unilatérale : La longévité d’un organisme pluricellulaire dépasse celle
d’un unicellulaire. En ce sens l’hérédité pluricellulaire est un progrès.
Cependant c’est l’hérédité cellulaire qui sous tend l’hérédité pluricellulaire.
La cellule est plus éphémère que l’organisme pluricellulaire, mais les
« mécanismes » conservateurs du niveau cellulaire survivent à ce dernier
(même s’ils évoluent aussi) [voir la conclusion du chapitre précédent].
Il est temps de revenir sur la position à adopter vis à vis de la
théorie mendélienne de l’hérédité. Mendel a découvert des lois de ségrégation
indépendante de facteurs (les gènes) de génération en génération, contre le
principe d’hérédité-fusion communément admise à son époque. Son mérite en a été
de découvrir l’existence objective des gènes (celui de Morgan, à sa suite, fut
celui de découvrir à travers les indépendances et les liaisons génétiques
héréditaires, l’existence du lien matériel entre gènes et chromosomes –ou
groupes de gènes liés).
Mendel, Morgan, les généticiens du développement actuels n’ont jamais
travaillé que sur la transmission de caractères récessifs non adaptatifs voire
délétères, c’est à dire des « non-caractères », des absences de
caractères sauvages[22].
Entre l’existence de gènes particulaires dont la destruction retentit sur le
phénotype sous forme de variations diverses et (retournement de l’analyse)
l’existence d’un phénotype global en tant que somme discrète de caractères
génétiques (les caractères ne fusionnent pas parce que les gènes ne fusionnent
pas), le fossé est vite franchi, pour peu qu’on ignore la distinction
dialectique du général et du particulier. Les gènes existent objectivement,
mais le principe d’hérédité-fusion[23]
aussi !
C’est en ce sens que le mendélo-morganisme doit être attaqué. Le
phénotype global résulte d’interactions multiples : synthèse protéique
dominée par des gènes, actions réciproques entre gènes (selon leurs proportions
respectives et plus encore, semble t-il, leurs positions relatives),
interactions ADN – ARN – polypeptides immatures – protéines, contribution
collective des activités moléculaires, cellulaires et environnementales dans la
réalisation des propriétés, des structures générales de l’organisme.
b. Le statut de la sélection naturelle et la question de l’ectogenèse
D’un côté les mutationnistes partisans d’une évolution aveugle et
accidentelle des êtres vivants (autogenèse), de l’autre, les sélectionnistes
affirmant que c’est le « milieu » qui les « sculpte » et
les transforme (ectogenèse). Avons nous à faire à deux partis scientifiques en
conflit, l’erreur et la vérité, ou à une querelle de salon entre métaphysiciens
acharnés ? Voilà quasiment toute l’histoire de la science de l’évolution…
Ce débat poussa par exemple Filiptchenko lui même, savant soviétique du début
du vingtième siècle, à distinguer en chaque espèce des caractères
« organisationnels » d’une part, des caractères
« adaptatifs » d’autre part : Une telle distinction, intolérable
du point de vue marxiste, a néanmoins l’avantage d’expliciter la teneur d’un
débat encore très actuel :
Un même milieu porte de nombreuses espèces distinctes les unes des
autres. Un milieu ne sculpte donc pas une espèce idéale qui lui serait
spécifiquement adaptée (ectogenèse) : Il est habité par des types
préexistants tous au niveau de la biosphère mais localement transformés… Le
type « oiseau » est représenté par des échassiers sur un marécage,
par des manchots sur la banquise, etc. Les « caractères
organisationnels » propres à l’oiseau persistent, leur « caractères
adaptatifs » (miniaturisation du plumage chez le manchot, allongement des
pattes et du bec chez les échassiers, …) varient.
Mais le problème reste entier : Ces deux types de caractères
restent physiquement indissociables. Le plumage du manchot est un caractère
organisationnel (participe à l’homéothermie propre aux oiseaux) et adaptatif
(miniaturisé, il en devient hydrodynamique). Jamais nous ne sortons du problème
singulier de la téléologie…
Contre la téléologie, un exemple parmi tant d’autres : Considérons
un Homme adulte. Ses lèvres contribuent directement à l’usage de la parole. Les
lèvres servent à la parole… Considérons l’Homme de plus loin : Grâce à la
science… nous découvrons bientôt qu’il a été petit avant d’être adulte, qu’il
existe bien d’autres mammifères dotés de lèvres, sans parler pour autant. En
effet, les lèvres assurent une fonction primordiale, bien que transitoire, chez
tous les mammifères ; celle de téter. Tirons une leçon de cet exemple.
L’existence des lèvres est à la fois nécessaire pour la tétée et fortuite pour
le langage. De tels exemples ne manquent pas dans la nature, et sont d’ailleurs
cause depuis plusieurs siècles d’une illusion fixiste selon laquelle chaque
être vivant est tellement bien adapté à son milieu qu’il ne pourrait pas
évoluer sans mourir (loi de corrélation des organes). En ce qui concerne les
lèvres, on ajoutera que c’est chez l’Homme qu’elles sont le plus mobile, en
rapport avec l’élocution (valeur adaptative) qui lui est spécifique. Posons
donc ceci :
-
Un caractère ne peut être considéré
seul : Sa valeur adaptative et fonctionnelle tient à sa coexistence avec
d’autres caractères (coadaptation) du même organisme (tous les autres organes
de la phonation), d’un autre organisme (mamelles de la mère), du milieu
physique[24] (consommation d’eau).
-
Un caractère est donc à la fois
organisationnel et adaptatif, cette distinction étant tendancielle. Le
développement adaptatif d’un caractère est déterminé (orienté) à partir de
plusieurs potentialités initiales : D’abord la tétée, ensuite la parole.
Ainsi, la fonction ne crée pas l’organe (contre Lamarck, figure de proue de
l’ectogenèse), mais l’organe crée la fonction (ou plutôt des fonctions).
Cela ne signifie nullement que l’organe n’est pas créé à son tour. Cela
ne signifie pas non plus que son perfectionnement est indépendant de la
fonction nouvelle qu’il accomplit. Quelques éléments d’explication avec
l’exemple des pluricellulaires…
Au niveau pluricellulaire, on soulignera les rapports entre quantité et
qualité ; Toute forme discrète d’un individu apparaît au terme d’un
développement conditionnant des paramètres quantitatifs. La plume du manchot
est une plume dont le développement cesse très tôt (par rapport au reste du
corps), le bec et les pattes de l’échassier sont des organes dont le
développement se poursuit anormalement (par rapport au reste du corps). Plus
que la taille, la forme de l’organe elle même résulte d’un développement
différentiel de ses parties (la courbure du bec du courlis, un échassier
dénichant des vers dans des terriers courbes, résulte de son accroissement plus
rapide en face dorsale qu’en face ventrale). En aucun cas on ne peut affirmer
que de tels caractères sont « virtuellement » présents dans les gènes
de la cellule-œuf. Ils ne sont présents nulle part avant leur fixation
apparente, et ne peuvent résulter que d’interactions auto-ectogénétique pendant
le développement général de l’individu. Pas de préformisme… Reprenons ici à
notre compte l’idée de Waddington selon laquelle le développement est un chemin
traversant un ensemble complexe de contraintes épigénétiques. Ce chemin fait
partie intégrante de l’hérédité pluricellulaire au même titre que l’hérédité
cellulaire (son niveau inférieur). Les contraintes (contradiction entre
tendances internes au
développement spatial et obstacles extérieurs)
explique les
développements différentiels de la morphologie d’un individu, en dehors de
toute téléologie. Chaque forme donne lieu a priori à une infinité de
potentialités fonctionnelles.
On connaît la capacité qu’a une bouture de néoformer des racines et des
feuilles, celle du lézard à régénérer partiellement la queue qu’il a perdu. De
telles capacités, montrant que le passage qualitatif cellules totipotentes –
cellules différenciées n’est pas absolument irréversible, même au stade adulte
(surtout chez les animaux primitifs ; les vers doués de scissiparité par
exemple), peuvent être rapprochées de celle qu’a la cellule-œuf de former un
organisme pluricellulaire entier. Un ensemble de cellules se développe dans
trois champs de contraintes :
1.
Les potentialités propres des
cellules (remarquons que l’environnement d’une cellule est son milieu
extracellulaire mais aussi l’état initialement hérité de son cytoplasme).
2.
L’environnement organique direct
(réserves extra-embryonnaires, reste de l’organisme, organisme maternel)
3.
L’environnement inorganique
extra-corporel.
Ces trois champs sont indissociables : Quel marxiste désignera l’un
d’eux à l’exclusion des autres, sous le terme de sélection naturelle ?
Néanmoins : Le premier champs correspond à l’hérédité cellulaire, ou
conservation de la totipotence équivalent au stade unicellulaire. Le second
correspond à l’hérédité pluricellulaire, ou reproduction d’un processus
complexe, amenant les cellules à des stades différenciés, et l’organisme à des
formes « achevées » accomplissant leurs fonctions. La seule réussite
de la conservation cellulaire est de laisser au moins quelques cellules
au stade totipotent ; les gamètes. Le premier et le second champs
consistent à donner à l’individu en développement un milieu
« stable » (reproductibilité), même si chacun de ces champs est
affecté d’instabilités à plus ou moins long terme. Le troisième champs n’est
pas le plus stable (évolutions climatologiques, géologiques, géographiques).
Les néodarwiniens l’appellent le « milieu », l’acteur d’une sélection
naturelle aveugle. Il a pourtant lui aussi, il faut le remarquer, une part de
stabilité : chez les animaux ovipares par exemple, le milieu du
développement des œufs est choisi par les parents sur de tels critères.
Ainsi le développement de l’organisme dépend d’une contradiction
dynamique entre conditions stables et conditions changeantes du milieu (trois
champs). Nombreuses sont les conditions changeantes de ce milieu qui ne sont
pas en interaction avec l’organisme : en particulier les conditions
climatiques et géologiques. L’évolution des espèces passe inévitablement par le
dénouement incessant de cette contradiction[25].
L’organe apparaissant avant la (les) fonction(s) historiquement (et
embryologiquement), la sélection naturelle ne peut orienter le phénotype
(organes fonctionnels) vers une meilleure adaptation. Nous considérerons
l’orientation sous un autre angle [voir annexe « Deux tabous :
Convergence et Orthogenèse »].
Quand, dans notre contradiction, les conditions changeantes deviennent
prépondérantes par rapport aux conditions stables, une réadaptation de l’espèce
devient nécessaire, au sens strict du mot. Ce que les néodarwiniens
opposent sous les termes mutagenèse et sélection naturelle, ne constituent
qu’une seule et même pression désorganisatrice, « anonyme »,
appliquée sans distinction sur deux niveaux structuraux à la fois.
Au niveau de l’organisme pluricellulaire, le milieu, qui met en
mouvement tout le processus ontogénétique, est remis en question :
L’individu, conservant les modalités de conservation cellulaire, réagit en
changeant (le moins possible et en rapport avec l’importance du stress). Les
changements sont non-orientés et très variés (voir la partie précédente sur
l’autogenèse). Telle est la mutagenèse[26].
Au niveau du groupe spécifique, le milieu changeant décime la population
(morts brutales ou baisse démographique par hausse de la mortalité et baisse de
la natalité). Toutes les formes deviennent minoritaires. La variation la plus
adaptative venant d’apparaître et minoritaire
comme les autres, deviendra de fait majoritaire en relativement
peu de générations, permettant seule le re-développement de l’espèce (alors
modifiée). Telle est la sélection naturelle.
Cette fusion conceptuelle mutagenèse / sélection naturelle apporte un
complément important à notre théorie : La contradiction conditions
stables / conditions changeantes est antagoniste :
-
Les conditions changeantes finissent
toujours par dominer les conditions stables.
-
Cette prépondérance crée une
accumulation quantitative de modifications structurales chez les individus
d’une population menacée par le changement, toutes minoritaires, donc
incapables de conquérir « nécessairement » la population entière (en
déclin). Une forme avantageuse ultra-minoritaire dans une population très
nombreuse, ne se répandra en effet dans cette population qu’après une quantité
incalculable de générations : Plus le temps de cette conquête est long,
plus elle est en proie à des évènements extérieurs fortuits.
-
En décimant la population, la forme
avantageuse ne sera plus minoritaire et deviendra ultra-majoritaire en peu de
générations : C’est un saut qualitatif nécessaire.
Marx et Engels acceptaient le couple hasard-sélection comme explication
provisoire de la spéciation. Provisoire, cette formule l’était en effet :
Nous n’avons pas nié les variations inter-individuelles –ou polymorphisme-,
celles-ci n’échappent pas à la causalité, elles résultent au contraire des
modalités mêmes de la conservation du groupe spécifique (espèce). Nous ne nions
pas non plus la sélection naturelle ; Nous indiquons simplement qu’il existe
un conditionnement réciproque direct entre celle-ci et le polymorphisme
(variation inter-individuelle) : Ce n’est pas le « milieu
extra-corporel » mais l’« inadéquation espèce-milieu » -ou stress-
qui effectue la sélection naturelle, non pas par un simple tri des individus
les meilleurs mais par un processus nécessaire qui en décimant quantitativement
la population finit par accélérer l’extension des nouvelles formes
individuelles les plus adaptées (réponses individuelles non
« préméditées » à ce stress, c’est à dire à l’inadaptation) à
la micro-population (nouvelle espèce).
Notons enfin, et l’enjeu est important, qu’il existe une lutte
intra-spécifique dans le cadre du saut qualitatif parmi les
« survivants » de l’espèce. Cette « lutte du plus apte pour sa
survie » pour employer les termes de la science bourgeoise, magnifiée par
les néodarwiniens comme une loi naturelle, n’est bien sûr qu’une loi relative à
un contexte donné[27].
S’adressant à Friedrich Lange, sociologue et philosophe allemand membre
de la première internationale, qui soulignait le caractère bourgeois des
concepts darwiniens, notamment de la loi de la sélection naturelle, Engels fit
la remarque suivante :
« Moi aussi
j’ai été frappé, à la première lecture de Darwin, par la ressemblance frappante
entre sa présentation de la vie végétale et animale et la théorie de Malthus.
Seulement j’en ai tiré une autre conclusion que vous, savoir : que ce
qu’il y a de moins glorieux dans le développement bourgeois contemporain, c’est
qu’il n’a pas encore dépassé le niveau des formes économiques du règne animal.
Pour nous, ce qu’on appelle les « lois économiques » ne sont pas des
lois éternelles de la nature, mais des lois historiques, qui naissent et
disparaissent, et le code de l’économie politique moderne, dans la mesure où
l’économie l’établit vraiment de façon objective, n’est pour nous que le résumé
de l’ensemble des lois et des conditions qui seules permettent à la société
bourgeoise moderne de continuer d’exister, en un mot : l’expression
abstraite et le résumé de ses conditions de production et
d’échange. »
F.Engels. Lettre à F.Lange, 29 mars 1865.
c. Un modèle simplifié du processus de spéciation
Correction mutation
Barrière gène-prot
Fig.8 ;
Mécanisme général d’une spéciation. M ; milieu initial, M’ ;
milieu final, SN ; sélection naturelle, Quant ; modification
quantitative, qual ; modification qualitative, W ; travail des
organismes sur leur milieu commun, EG ; effet de groupe (stabilisateur),
d ; décimation, e ; expansion, Type A ; population polymorphe
autour de l’archétype A, Type B ; population polymorphe autour de
l’archétype B. Les graphiques ne rendent pas compte de l’irréversibilité de la
transformation A B (Loi de Dollo, réhabilitée par Gould)
Nous prendrons volontairement la spéciation sous son cadre le plus
général, en tant que processus marqué par des étapes schématiques,
« fixées » pour l’occasion. Une telle description (fig.8) se destine
à identifier chaque aspect de la transformation sous sa forme la plus simple et
la plus exagérée ; il faut toutefois garder à l’esprit le caractère
complexe du processus général et conséquemment le caractère particulier et
unique de chaque exemple de spéciation. Ces aspects sont à tel point
inter-dépendants et mouvants qu’une formule exhaustive purement mécanique de la
spéciation ne saurait exister aux yeux d’un dialecticien.
Suivons les étapes principales de la transformation d’une espèce A en
une espèce B (qu’elle soit micro ou macro-évolutive) :
a. L’espèce A occupe un milieu M (milieu physique, espèces prédatrices,
ressources organiques et minérales, constructions de l’espèce A) qui contrarie
peu son développement quantitatif et qu’elle modifie peu : apparente
stabilité résultant d’une situation passée au cours de laquelle l’activité
assimilatrice A→M et l’action sélective M→A ont façonné une
co-adaptation A↔M optimale. Le polymorphisme de l’espèce s’en trouve
particulièrement restreint, organisant celle ci autour d’un type moyen
sur-représenté, nommé « type A ».
b. Ce milieu vient à changer qualitativement, de façon contingente
(rencontre brutale d’une série causale indépendante de A↔M) ou nécessaire
(situation où l’équilibre adaptatif relatif est impossible du fait des
caractéristiques propres à A ou à M : l’accroissement progressif de ce
déséquilibre A→M > M→A [surpeuplement et excès consécutif de
transformation du milieu] ou A→M < M→A [adaptation trop
spécialisée, polymorphisme trop affaibli, donc trop fragile vis à vis des
moindres fluctuations environnementales] suscite une inadaptation brutale).
c. Dans le cas où cette brusque inadaptation ne provoque pas
l’extinction totale de l’espèce A, celle-ci subit au moins une décimation forte
(modification quantitative). Chaque individu de la population souffre alors
d’un stress causé indissociablement par l’inadaptation A│M’ elle
même et par l’amoindrissement de l’« effet de groupe » (dont
l’activité a normalement pour effet d’atténuer cette inadaptation).
d. Le stress retentit à tous les niveaux d’intégration de l’espèce A,
détruisant partiellement les processus compensateurs de celle ci : mécanismes
correcteurs (α) préservant le génome des mutations, barrière
ADN-Protéines (β) protégeant le génome de la rétro-action des produits
de gènes, barrière soma-germen (γ) protégeant les cellules
germinales de l’influence somatique, et barrière reproductive (δ)
empêchant à la fois l’hybridation inter-spécifique (homogénéisante) et
l’« auto-fécondation » (hétérogénéisante), ne fonctionnent plus
correctement.
e. A toutes les échelles, la variabilité explose, non directionnelle aux
échelons inférieurs, mais directionnelle aux échelons supérieurs : l’ADN
subit de profonds remaniements, intra et inter-géniques, aléatoires et
abondants (α), une rétro-action des produits inactifs en excès ou sur-actifs
en carence sur une partie ponctuelle du génome cible nécessairement les zones
génétiques les plus directement corrélées à l’inadaptation A│M’ sans
toutefois diriger leurs transformations (β), spatialement ces
modifications seront préférentiellement (mais pas seulement) localisées au
niveau germinal (γ), enfin, la typologie qui prévalait pour l’espèce A
n’empêche plus des individus A1, A2, Ax, B ou
C issus de A, même en uniques exemplaires (qui auraient été condamnés à rester
sans descendance avec la barrière d’espèce), d’être interféconds
(δ) : Les transformations sont donc extrêmement variées et toutes
potentiellement transmissibles aux générations suivantes.
f. Chaque mutant deviendra d’autant plus vite majoritaire que, d’une
part, le nombre d’individus décroît et que, d’autre part, la stratégie de
reproduction sexuée s’intensifie, traversant le crible de la sélection
naturelle. Celle ci n’est plus alors (seulement) déséquilibrante, mais (aussi)
stabilisante : les formes nouvelles subissent cette sélection au niveau
moléculaire (la reconstitution d’un nouveau génome doit répondre aux nombreuses
contraintes de l’infrastructure cellulaire, que celles ci aient
superficiellement ou profondément changé), au niveau cellulaire (tri des
embryons viables) et au niveau de l’organisme (sélection des individus selon
leur valeur adaptative).
g. On passe donc d’un changement quantitatif (décimation et
conséquemment excès de ressources dans le milieu dépeuplé) à un changement
qualitatif (prépondérance d’autant plus rapide d’une forme co-adaptée
B↔M’ dans la population A’ que celle ci se dépeuple en forme A), sous
l’influence d’un nouveau milieu M’ lui même re-stabilisé par l’activité de B,
formant bientôt le « type B ». Les barrières multipolaires s’étant
progressivement reconstituées sous l’effet de la pression sélective
stabilisante de M’, les formes hégémoniques B constituent à cette occasion une
nouvelle espèce, qualitativement (typologiquement) distincte de l’ancienne
espèce A.
C. Conclusion
Les uns auront tendance à ne voir dans cette analyse qu’une opposition
systématique aux thèses indéterministes à la mode, les autres au contraire un
retour honteux au « réductionnisme » d’antan.
Notre essai s’inscrit pourtant en toute légitimité dans la crise
actuelle du néodarwinisme, et ce pour deux raisons au moins. La première tient
à la pratique scientifique elle même : bornée comme nous l’avons vu entre
le holisme d’avant-garde et le déterminisme d’« arrière garde », elle
s’oriente provisoirement mais nécessairement vers des conceptions dialectiques
inédites et prometteuses, offrant malgré elle à notre analyse l’opportunité de
nombreuses découvertes renvoyant dos à dos réductionnisme et indéterminisme. La
deuxième renvoie aux contradictions qui enferment l’idéologie officielle dans
un rejet de toute synthèse dialectique explicite : une telle synthèse la
précipiterait à coup sûr dans l’impasse qu’elle cherche inconsciemment et
vainement à éviter, voire à taire.
Arguments issus
de la pratique
L’étude de la lignée humaine, c’est à dire des mécanismes qui ont
historiquement donné naissance à notre espèce, devient apparemment l’axe
d’avant garde des sciences de l’évolution, l’exemple phare qui influence la
théorie évolutionniste dans son ensemble.
Or la paléoanthropologie vacille aujourd’hui, pour des raisons multiples
et indépendantes. Profitons de ce contexte et citons en trois, dont la portée
dépasse objectivement le cadre de la seule lignée humaine.
α. Sur l’arbre de l’évolution, on s’accorde
à définir la lignée humaine comme l’ensemble des fossiles qui sont plus
apparentés morphologiquement à l’Homme moderne qu’au chimpanzé (le chimpanzé
étant l’espèce actuelle qui nous est la plus proche génétiquement et
morphologiquement). Ainsi l’australopithèque, pourtant très simiesque, est plus
étroitement apparenté à l’Homme qu’au chimpanzé parce qu’il est bipède
permanent (libération des bras et conséquemment développement de l’intelligence
par la conception et la manipulation d’outils). A partir de l’ancêtre commun
théorique à l’Homme et au chimpanzé, l’émergence accidentelle de la bipédie
permanente sous tendrait le « buissonnement » ultérieur de la lignée
humaine. Bipédie accidentelle et développement non linéaire de la lignée sont
les bases de l’indéterminisme actuel dans les conceptions anthropologiques.
Or une découverte fort irritante commence à se faire écho dans la presse
scientifique ; les plus anciens bipèdes permanents connus à ce jour, Tumaï
et Orrorin sont datés respectivement à –7 et –6 millions d’années, grâce à des
méthodes de plus en plus précises et rigoureuses. Selon des calculs de plus en
plus exacts eux aussi, relatifs aux comparaisons entre les ADN d’Homme et de
chimpanzé actuel, l’ancêtre commun théorique Homme/chimpanzé, qui n’est donc
pas encore bipède permanent, ne peut être antérieur à –5 millions
d’années !
Première conséquence ; Tumaï et Orrorin n’appartiennent pas à la
lignée humaine et furent donc des « impasses évolutives ». Deuxième
conséquence ; la bipédie permanente, qu’on postulait accidentelle, est
donc apparue plusieurs fois dans l’histoire évolutive des primates. Troisième
conséquence ; si la bipédie est apparue plusieurs fois, elle n’est donc
pas accidentelle… elle était au contraire nécessaire ! Autrement dit, tout
nous pousse à considérer que si l’Homme n’était pas apparu, d’autres espèces
bipèdes permanents et intelligentes auraient émergées à sa place !
En fait ce type de découverte n’est pas nouveau ; celle ci est
simplement beaucoup plus précise parce que plus récente. On sait par exemple
que de nombreuses familles de poissons sans aucun lien de parenté entre elles,
sans parenté non plus avec les premiers tétrapodes (batraciens) ont acquis des
poumons fonctionnels en plus de leurs branchies (plus ou moins régressées) ou
des structures analogues rendant possible la vie hors de l’eau. Autre
exemple ; l’Archéoptéryx, petit « dinosaure » doté de plumes et
d’ailes, n’est pas apparenté aux premiers oiseaux (dont le bassin est de type
ornithischiens) qui lui sont pourtant contemporains. Autre exemple encore ;
la classe des batraciens n’a pas un seul ancêtre commun, mais au moins deux,
malgré son apparente homogénéité morphologique.
Autant de transformations qui semblent donc « nécessaires »,
c’est à dire non accidentelles… Autant d’affronts à l’arrogance de l’avant
garde indéterministe !
β. Le perfectionnement des méthodes de
datation commence de plus à relativiser le modèle buissonnant de la lignée
humaine, au profit d’un arbre beaucoup plus complexe de
séparations-hybridations entre espèces proches. C’est le modèle
« tokogénétique ».
Le dogme typologique est donc aujourd’hui en sérieuse difficulté ;
rappelons que si des hybridations naturelles sont possibles entre espèces
distinctes, cela veut dire que 1) les barrières d’espèces sont des processus
objectifs autant que « contournables » et admettent donc
dialectiquement leur contraire, 2) Mitchourine et Lyssenko avaient
raison !
γ. L’Homme et le chimpanzé ont des génomes
homologues à plus de 99%. Pire ; la plupart de leurs gènes le sont
carrément à 100%. De plus, une différence de 1% étant bien sûr relative à la
quantité de gènes du génome, on rétorquait à l’inacceptable découverte que 1%
d’un très grand nombre de gènes représente malgré tout une différence non
négligeable : Or ce nombre, déterminé récemment, est finalement
particulièrement réduit (l’Homme possède par exemple autant de gènes qu’une
simple mouche)
Découverte complémentaire : Ce 1% de différence réside
essentiellement dans des séquences spéciales souvent situées hors des gènes
(entre les gènes ou dans les introns) et impliqués dans les transpositions
génétiques c’est à dire la duplication (quantitative) de certains gènes en
fonction des besoins ponctuels et la réorganisation spatiale (qualitative) des
gènes entre eux.
La distinction évolutive Homme/chimpanzé concerne donc moins
l’accumulation de mutations ponctuelles accidentelles dans les gènes, ce
« moteur de l’évolution » qui fait aujourd’hui encore l’unanimité,
que la réorganisation indissociablement quantitative et qualitative des gènes
entre eux.
Preuve est donc faite que des mécanismes compensateurs perfectionnés
(ici les transpositions) dominent les mutations ponctuelles (base du
néodarwinisme) dans les processus moteurs de l’évolution biologique. Autrement
dit, le couple hasard/sélection d’apparence indéterministe est donc en fait un
mode évolutif englobé dans un processus plus large qui lui est contraire ;
la propriété (vainement) auto-conservatrice de la matière vivante.
Précisons ici qu’une expérience récente (citée dans La Recherche
377, juillet-août 2004), « révolutionnaire » mais si peu médiatisée
dans sa signification théorique, donne à notre modèle un poids inespéré :
Lorsqu’une souris mâle est soumise à un air pollué (stress), sa « lignée
germinale » (cellules qui se divisent le plus intensément dans son
organisme, pour produire des spermatozoïdes en permanence, et qui subissent
donc potentiellement le plus de mutations) mute sélectivement sur des
« points chauds » (sic) de l’ADN. Ces points chauds ne se tiennent
pas au niveau des gènes, mais précisément sur des séquences non codantes
dirigeant duplications et recombinaisons spatiales des gènes dans le noyau.
Faut-il préciser qu’affectant préférentiellement la « lignée
germinale », et ce de façon sélective mais non orientée (l’ADN ne mute pas
aveuglément mais sur des portions précises, cependant que les mutations elles
mêmes sont aveugles : c’est la très grande quantité des spermatozoïdes
produits qui rend possible l’élection d’une recombinaison non seulement viable
mais aussi meilleure), ces mutations sont de fait héréditaire.
De quelques théories fondées sur la pratique actuelle, hétérodoxes mais
aussi très ponctuelles et cloisonnées, peuvent germer des développements
encourageants de notre synthèse dialectique : Théorie tokogénétique
(Henning, T.Holliday) sur les hybridations évolutives, théorie des
« niches morphologiques » (C.Morris)[28]
affirmant que les morphologies compatibles avec la vie ne sont pas infinies et
que seules quelques unes conviennent à tel ou tel milieu (tentative
d’explication des processus de convergence évolutive), théorie des transitions
majeures (JM.Smith) qui argumente impeccablement la complexification croissante
du vivant et tente une modélisation dialectique égoïsme/coopération des
transitions historiques entre niveaux d’organisation du vivant
(uni/pluricellulaires, auto-organisation des systèmes mutualistes en général).
Autant d’arguments contre les indéterministes incurablement allergiques aux
notions de complexification croissante et de nécessité évolutive.
Synthèse
incompatible avec l’idéologie
Notre synthèse se fonde à la fois sur le néodarwinisme finissant et sur
les théories alternatives plus ou moins implicites d’aujourd’hui. Elle n’est
donc concrètement possible qu’aujourd’hui. Cependant elle est aussi plus
inacceptable pour la biologie actuelle qu’elle ne l’aurait été il y a trente
ans, à l’apogée du néodarwinisme (exception faite bien sûr de notre étude du
lyssenkisme). La raison en est simple ; elle se réfère au néodarwinisme, à
la génétique moléculaire autant qu’aux thèses antagonistes de Kupiec et Sonigo.
Mieux ; elle ne se rapporte finalement ni à l’une ni à l’autre, chacun des
camps se construisant isolément en opposition métaphysique à l’autre, c’est à
dire contre toute synthèse.
Ainsi l’hérédité cellulaire est bien conçue comme l’émergence de
processus auto-réplicatifs dont le perfectionnement, l’extrême raffinement,
résultent à l’origine d’un pur hasard/sélection à l’échelle moléculaire. Cette
hérédité cellulaire procède donc comme le prévoit le néodarwinisme, à ceci prés
que la vertu auto-réplicative des acides nucléiques précède le processus
darwinien de hasard/sélection. Ce dernier ayant progressivement mis en place
des mécanismes conservateurs de plus en plus perfectionnés, ils ont devancé
puis dominé le processus hasard/sélection (saut qualitatif) dans tous les
développement de l’histoire de la vie. L’évolution fut dés lors gouvernée par
les contradictions internes à ces mécanismes conservateurs eux mêmes.
Cette histoire de la vie n’est pas pour autant devenue anti-darwiniste.
Le processus hasard/sélection, partiellement dominé dans les termes que nous
avons précisé plus haut par les mécanismes conservateurs, est toujours préservé
comme une stratégie privilégiée d’évolution (ce n’est pas la seule) bien
qu’inféodée à la propriété conservatrice fondamentale de la matière vivante. Le
processus de « conservation des propriétés conservatrices » interrogé
dans le chapitre précédent répond donc à la loi dialectique bien connue et
combien polémique (chez les maoïstes en particulier) de négation de la
négation…
A ce stade très « finaliste », ce sont les sonigiens et autres
indéterministes qui grimassent de dégoût. Pourtant l’hérédité pluricellulaire,
fondée sur un renversement dialectique non total des processus de l’hérédité
strictement cellulaire, devraient plaire à ces derniers ; elle se fonde
sur une négation de tous les dogmes de la génétique moléculaire (distinction
soma-germen, barrière d’espèce, stéréospécificité moléculaire). Malheureusement
cette négation finaliste des dogmes n’est qu’en apparence analogue à leur
négation indéterministe.
Voilà donc une théorie sans camp, construite à l’heure où les camps
n’ont plus de théorie (c’est à dire de synthèse théorique réelle) !