Chapitre 5 : Niveaux structuraux
et stades de l’histoire naturelle
Théorie dialectique de l’évolution
«Pour savoir de façon réellement complète ce que c’est
que la vie, il nous faut passer en revue toutes les formes sous lesquelles elle
se manifeste, de la plus inférieure à la plus élevée »
«La science de la nature en est actuellement
arrivée au point de ne plus pouvoir échapper à la synthèse dialectique »
Engels (Anti-Dühring)
On retrouve les lois de la dialectique de façon dispersée à travers les « camps » du champ
de bataille scientifique, mais cette « synthèse dialectique »
qu’Engels appelait de ses vœux ne semble pas imminente. A ce stade, constatons
simplement que, si le matérialisme est la posture nécessaire des savants dans
l’exercice de leur pratique scientifique, l’approche dialectique semble de
surcroît fonctionner dans cette pratique, puisqu’elle émerge – certes
inconsciemment – au moins spontanément à plusieurs niveaux contre les courants
plus ou moins raffinés de l’idéalisme néodarwinien. Tentons donc à présent
d’élaborer notre synthèse. Nous disposons à cette fin, 1) des thèses philosophiques du matérialisme dialectique, 2) des données
contemporaines de la biologie.
Thèse
1 : La matière existe objectivement.
Thèse 2 : On ne peut réduire la matière à une de
ses formes (Holisme).
Thèse
3 : La matière est « une » (Monisme) : Il n’y a qu’un
monde.
Thèse
4 : La qualité est ce qui distingue une chose d’une autre.
Thèse 5 : La vie répond à un seul principe
organisateur ; la matière (anti-vitalisme).
Loi dialectique
A (Mouvement dialectique) :
Thèse 6.a : La manière d’être de la matière est
le mouvement.
Thèse 6.b : Ce mouvement auto-dynamique
nécessaire ne peut être qu’accéléré ou ralenti.
Thèse 6.c : Il est un processus dirigé
( non-cyclique).
Loi dialectique
B (Actions réciproques) :
Thèse 7.a : L’univers est un tout complexe
organisé.
Thèse 7.b : Ses aspects sont interdépendants.
Thèse
7.c : On entend par mouvement nécessaire celui qui est l’effet
d’une cause.
Loi dialectique
C (Contradiction) :
Thèse 8.a : La contradiction est la lutte et
l’unité des contraires.
Thèse
8.b : Un mouvement auto-dynamique a pour moteur(s) une (des)
contradiction(s) principale / secondaire(s).
Thèse
8.c : Une contradiction peut être antagoniste [thèse 9.b] ou non-antagoniste.
Loi dialectique
D (Quantité/Qualité) :
Thèse 9.a : Le mouvement transforme la matière
de l’inférieur au supérieur, du simple au complexe.
Thèse 9.b : Une contradiction antagoniste
provoque au terme d’une accumulation quantitative, un saut qualitatif,
dépassement de cette accumulation.
Rappelons pour éviter certains contresens que les
anticommunistes instrumentalisent si complaisamment, que cette synthèse n’a pas
la prétention de cristalliser la recherche pour le reste des temps; elle se
limitera au contraire à deux niveaux : 1) Au niveau pratique, il s’agit
d’une construction strictement fondée sur les données actuelles, donc sujette à
modifications, améliorations et corrections[1].
2) Au niveau théorique, cette construction est un instrument dirigé contre
l’idéalisme, destiné à le déloger des positions qu’il occupe actuellement sur
la question de l’évolution biologique…L’exposition des thèses matérialistes
dialectiques nécessaires à cette étude servira de base conceptuelle préalable : Leur efficacité sera évaluée pas à pas au
cours de la construction –l’étude n’ayant pas non plus pour but de
« prouver » rétrospectivement ces thèses-.
Une définition de la vie requiert l’éclaircissement de l’apparente
opposition entre les thèses 2 et 4 dans le contexte de notre étude :
L’être vivant est manifestement une chose qui se distingue qualitativement de
son environnement [thèse 4], mais l’être vivant et son environnement sont
indissociables [thèse 2], la preuve en est que l’être vivant est une structure
matérielle qui renouvelle en permanence ses molécules constitutives (fonction
de nutrition) à partir de la matière environnante, tandis que l’environnement
est lui même transformé par l’être vivant. Pas d’opposition, à condition de qualifier
ce qui distingue ces deux matières. L’expression matière vivante n’aura
pas un sens vitaliste (matière d’essence différente du reste de la matière)
mais visera justement à ne pas désigner la vie sous le terme structure
organisée avant d’en avoir fourni une définition argumentée. S’il est
permis de distinguer des qualités dans la matière, ne serait-ce que pour
montrer les interactions nécessaires entre ces qualités [thèse 7.b],
distinguons matière vivante et matière non-vivante et tentons une
définition claire de leur distinction.
D’abord, que la matière vivante soit une structure organisée, complexe
est une évidence : Encore faut-il préciser la légitimité des termes
« organisé » ou « complexe ». Ces qualités ne sont pas
spécifiques à la vie (pour un matérialiste évidemment[2]).
Le mouvement, qualité fondamentale de la matière [thèse 6.a] est la manière
d’être de toute matière, vivante ou non. Jusqu’ici, aucune distinction
dialectique apparente.
Venons-en donc à ce qui distingue spécifiquement la matière
vivante de la matière non vivante : C’est une évidence, un être vivant
renouvelle sa matière en permanence sans changer sa qualité d’être vivant
(jusqu’à un certain point) ;
Ce qui distingue la matière vivante de la matière non vivante, c’est la
tendance qu’elle manifeste à conserver sa
structure dans le temps.
Opposition apparente : Si la matière vivante est
« stable », dans sa structure, elle déroge à la thèse 6.a, thèse
fondamentale de la dialectique. Une explication s’impose : La vie répond à
un seul principe organisateur, la matière [thèse 5]. Sa matière est organisée
[thèse 7.a]. Juxtaposons la proposition irritante et la thèse 6.a : Le
mouvement interne de la matière vivante modifie nécessairement sa structure.
Cette désorganisation nécessaire peut-être freinée [thèse 6.b] par un
« processus conservateur »
apparaissant donc comme
une ré-action, un mouvement s’opposant, corrigeant
en permanence le mouvement désorganisateur. Ceci ne contrarie plus la
thèse 6.a. Remarque : il n’est pas dit ici qu’un mouvement désorganisateur
est un mouvement « destructeur », au sens où on l’entend
généralement ; Voici donc une proposition non-finaliste, non-mécaniste,
n’orientant pas plus la désorganisation vers la destruction que vers la
construction.
Pour être tout à fait clair et éviter d’ouvrir le flanc au vitalisme,
nous dirons que, 1) ce qui dans la matière en mouvement est doté d’une
propriété supplémentaire[3]
qui est de tendre à une « conservation » structurale sera appelé
« matière vivante »[4],
2) La question de l’origine de cette propriété doit être traitée dans le cadre
de la thermodynamique, émanant du principe de néguentropie qui établie la
possibilité d’organisation spontanée d’un système au détriment du reste de
l’univers conditionné par le principe entropique : N’est-il pas finaliste
d’affirmer que ce mouvement, étendu potentiellement à toute la matière,
produise toutes les structures possibles sauf celle qui aurait pour
propriété particulière de se conserver activement ? 3) Cette tendance à
conserver sa propre structure dans le temps, dans la mesure où elle aboutit –et
nous sommes forcés de constater qu’elle a objectivement abouti [thèse 1]-,
conduit à la nécessité d’une histoire de la matière vivante ; l’histoire
de la lutte pour l’existence.
Commençons donc l’exposé par un postulat simple issu d’observations
universellement reconnues ; La forme de vie la plus simple[5] est la cellule
[thèse 9.a]. La cellule sera considérée distinctement de son environnement quel
qu’il soit ; eau, air, reste d’un
organisme pluricellulaire, etc., de façon à mieux cerner leurs
interactions : D’un côté la matière environnante, en mouvement,
« agissant » par son mouvement sur la cellule de l’extérieur, et
transformée plus ou moins intensément par cette cellule, de l’autre la cellule
elle-même, organisée, en mouvement : « luttant » contre le
mouvement externe et le mouvement interne à la fois, suivant la proposition
formulée précédemment.
A. Premier niveau structural, la cellule
Que fait donc une cellule pour elle même, si ce n’est de renouveler sa
matière en permanence ? De notre point de vue d’organismes complexes peu
de choses ; quelques « stratégies de lutte » contre la
déstructuration, découvertes très récemment et qui méritent qu’on s’y attarde.
Tout d’abord, bien sûr, une cellule est mortelle. C’est l’inéluctable
fin du mouvement désorganisateur de la matière vivante [thèse 6.b]. Toutefois,
la division cellulaire, encore appelée reproduction conforme – ou mitose- parce
que la cellule produit deux cellules dont les caractéristiques sont celles de
la première, peut être considérée comme une forme de lutte contre la mort
cellulaire (négation), en faveur d’une conservation « dans le temps »
de la structure cellulaire (négation de la négation).
De plus, à chaque division cellulaire, les gènes, qui doivent être
dupliqués avant d’être distribués à l’identique aux deux cellules-filles,
subissent d’inévitables erreurs de copie [thèse 6.a], les fameuses mutations.
On sait aujourd’hui que toutes les cellules sont dotées de systèmes
moléculaires « correcteurs » ; ce sont les systèmes SOS, SRM[6](enzymes
corrigeant les mutations, découvertes chez les bactéries puis chez tous les
Eucaryotes) et HSP906 (protéines de choc thermique réprimant
l’expression des gènes mutés sauf en cas de stress, découverte chez la
drosophile et généralisable).
On sait que ces systèmes s’ébranlent plus ou moins lors d’un stress
(élévation de température, radiations, toxicité, carence nutritive) accélérant
le rythme des mutations contre lesquelles ils sont censés lutter [négativement
(SRM) mais aussi positivement (SOS)]. En cas de « coup dur », il vaut
mieux en effet favoriser la diversité fonctionnelle et ainsi augmenter la
chance pour quelques cellules de survivre, et pérenniser au moins partiellement
la structure cellulaire préexistante.
Tout aussi intéressant, le couple génétique p53 / MDM26
fonctionne sur la base d’une étonnante contradiction : p53 agit en
réparant les mutations ponctuelles et en empêchant pendant ce temps les
divisions cellulaires. MDM2 agit au contraire en stimulant les divisions et,
c’est prouvé chez l’Homme, en favorisant la prolifération cancéreuse
(désorganisation partielle). C’est l’activité de p53 qui stimule
collatéralement la production de MDM2, celle-ci inhibant en retour la
production de p53 (régulation dynamique par rétroaction).
Lorsqu’une cellule dispose d’un « gène de résistance » contre
une toxine, la présence de celle-ci déclenche une duplication massive du gène
dans le noyau. Ceci a pour effet d’augmenter considérablement l’intensité de
son expression : Les duplicatas sont appelés « transposons »[7].
De telles transpositions sont en définitive très répandues dans les processus
cellulaires, et la recherche ne fait aujourd’hui que commencer à en déterminer
les causes et les modes d’action (chez une espèce diploïde, c’est à dire qui
possède deux versions, maternelle et paternelle, de chaque gène, la mutation
délétère d’une version peut être compensée par l’amplification –ou duplication-
de son homologue).
Si le fait qu’une cellule « oriente » la duplication massive
d’un gène ponctuellement nécessaire à sa survie ne choque personne, pourquoi
continuer à nier dogmatiquement la possibilité d’une « orientation »
de la mutabilité sur le ou les gènes rendus ponctuellement inutiles ou
délétères ?
Ne compliquons pas davantage… Nous avons cinq processus. Deux
d’entre eux sont des processus désorganisateurs internes ; la mort
cellulaire et la mutagenèse. Deux autres sont des processus de lutte
active ; la mitose et l’ensemble des mécanismes compensateurs. Il y a ici
dans les faits contradiction entre ces deux mouvements, opposés dans leur
direction par rapport à la structure cellulaire. Un dernier processus est celui
de l’influence désorganisatrice de l’environnement, inextricablement lié aux
précédents processus (fig.2).
Processus conservateurs et évolutifs liés à la cellule : Ve ; modifications du milieu ou variations environnementales,
Vi ; modifications internes, Ci ; constance infra-structurale,
Ce ; Constance externe ou ultra-structurale.
Fig.2
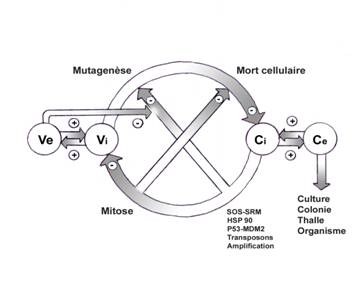
Première contradiction
Les mécanismes compensateurs, lorsqu’ils sont monopolisés par des
« contrariétés » extérieures, favorisent positivement les mutations
génétiques. Si on admet que des innovations génétiques permettent
potentiellement un supplément de résistance cellulaire aux contraintes
environnementales, il faut reconnaître que les mutations sont autant
désorganisatrices-destructrices que désorganisatrices-constructrices, dans le
sens où elles conduisent, au moins potentiellement, à l’acquisition de
nouvelles propriétés conservatrices : Pour
« créer » un nouveau
gène, il faut d’abord
qu’un ancien gène
se duplique, puis que le duplicata, d’abord inutile, accumule suffisamment de
mutations ponctuelles pour « devenir » le gène nouveau, assurant une
fonction nouvelle : C’est la théorie de la duplication génique.
Cette contradiction est antagoniste [thèses 8.a/8.c] : Systèmes
SOS-SRM, HSP90, P53/MDM2 et transposons agissent contre
les mutations génétiques
(ils les empêchent ou
les compensent) tout
en créant positivement les
conditions d’une innovation génétique lorsque le besoin s’en fait sentir. La
cellule n’a d’autre issue que d’évoluer, de changer, sous l’effet de cette
contradiction, et ceci parce qu’elle lutte pour la conservation de sa
structure, en cas de stress dangereux pour elle. On voit que la cellule,
« bien que vivante », ne déroge pas à la thèse 6.a, bien au
contraire.
On peut sans trop anticiper, mettre en avant la thèse 9.a : Sa
vocation étant de se conserver autant que faire se peut, elle évoluera
en conservant les caractères nouveaux dont l’accumulation la transforme
finalement !
Deuxième contradiction
On a choisi de présenter la division cellulaire comme une forme de lutte
contre la mort, pour la conservation dans le temps. Elle est une innovation
évolutive autant qu’un processus relatif à l’hérédité : Nous en
indiquerons ultérieurement les modalités. Ajoutons que ce processus a
collatéralement un avantage pour la conservation cellulaire .
La division cellulaire, considérée nécessairement comme une
prolifération cellulaire, produit des populations de cellules, susceptibles de
mener à bien une lutte « extérieure » de l’ensemble pour la
conservation de chacune (coopération cellulaire, bien connue des écologistes).
La coopération intercellulaire n’est plus un processus temporel mais un
processus spatial (division du travail, effet de groupe). La division produit
donc pour la cellule une superstructure (coopération cellulaire) favorisant de
l’extérieur sa conservation.
Par suite, la superstructure cellulaire associée à l’évolution
structurale cellulaire inéluctable, va permettre aux cellules de « s’auto
diriger » (pas de finalisme), toujours du seul fait de cette tendance
auto-conservatrice auto-dynamique (pas de vitalisme), vers une structure
nouvelle, plus complexe, pluricellulaire à spécialisation ; l’organisme.
La question de la conservation des propriétés conservatrices concerne
l’hérédité ; elle sera traitée dans le chapitre suivant.
Admettons ici que les caractères de spécialisation apparaissent par
innovations génétiques. Nous considérerons que l’organisme pluricellulaire est
une matière vivante plus complexe, susceptible d’assurer une conservation plus
efficace des cellules qui le composent. Du reste il existe de nombreuses formes
coloniales non spécialisées (un seul type de cellule pour tout
l’organisme : les thalles végétaux –algues, champignons, lichens- et
quelques animaux comme Volvox) qui attestent de la transition historique vie
unicellulaire / vie pluricellulaire. Ces colonies sont parfois composées de
cellules capables de retrouver plus ou moins leur autonomie dans certaines
conditions favorables (lichen, certains champignons, Volvox).
Pour conclure, les conditions extérieures accélèrent ou freinent
l’évolution cellulaire, mais celle-ci évolue en dernière instance par
auto-dynamisme, suivant des processus dirigés [thèse 6.c], tendanciellement
cycliques sans pouvoir l’être absolument.
-
Contradiction antagoniste secondaire : Pour lutter contre la désorganisation dans le temps, la cellule
prolifère (reproduction conforme), mais cette prolifération rend possible dans
l’espace le passage qualitatif du niveau structural de la cellule à celui de
l’organisme pluricellulaire ; passage qui est un résultat opposé à la
conservation structurale cellulaire (polymorphisme) [Thèse 9.a].
-
Contradiction antagoniste principale : Pour lutter contre la désorganisation génétique au cours du
temps, la cellule dispose de processus compensateurs, lesquels, en milieu
changeant (il l’est d’ailleurs par définition), finissent par accélérer une
évolution génétique [résultat opposé] sans laquelle elle ne pourrait survivre
[voir annexe « Variabilité génétique et structures compensatrices »].
B. Deuxième niveau structural, l’organisme
On objectera que les espèces pluricellulaires forment une minorité
quantitative et qualitative au sein du monde vivant, majoritairement composée
aujourd’hui d’espèces unicellulaires telles que les bactéries et les eucaryotes
unicellulaires. Cette marginalisation des espèces pluricellulaires, très
récente en science, argument favori des saltationnistes qui s’opposent
radicalement à la notion de complexification dans l’évolution et qui prétendent
qu’en définitive nous ne sommes jamais sorti de l’« ère des
bactéries », est évidemment anti-dialectique puisqu’ils considèrent –
c’est étonnant pour des évolutionnistes -
que si les unicellulaires sont restés majoritaires
en nombre d’espèces mais aussi en biomasse, c’est qu’ils ne passeront jamais à
l’avenir au stade pluricellulaire : Argument objectivement fixiste !
C’est nier le rôle des échelles de perception humaine du temps dans
l’approche de l’histoire évolutive (premier écueil), c’est également oublier la
nécessaire inégalité de développement des espèces actuelles (deuxième
écueil) : S’il faut un monde vivant dont toutes les espèces évoluent
ensemble au même rythme –alors que les interactions entre elles et avec leurs
milieux sont nécessairement hétérogènes- pour qu’un biologiste adhère à la
thèse de la complexification [thèse 9.a], il adhèrera du même coup à la thèse
peu matérialiste du miracle ![8]
Prenons donc l’organisme pluricellulaire comme nous avons pris la
cellule, tel qu’il se présente à nous, comme un « tout-complexe déjà
donné » selon l’expression consacrée : Tout d’abord, il est mortel.
Mais il n’est pas mortel parce que ses cellules sont mortelles (elles le sont)
mais parce qu’il est vaincu par la désorganisation de ses cellules. Ainsi il
n’est pas rare de voir survivre à un cadavre certaines de ses cellules (pour un
certain temps bien sûr), témoins par exemple, les cellules cancéreuses. De même
qu’on ne dira pas qu’une cellule meurt parce que ses molécules meurent, on ne
dira pas non plus que l’organisme meurt parce que ses cellules meurent, mais
parce que celles-ci ne fonctionnent plus ensemble (ne concourent plus à la
conservation structurale de l’organisme). On dira donc que la mort de certaines
cellules de l’organisme provoque la mort de celui-ci, que la mort de
l’organisme provoque à terme la mort de toutes ses cellules [thèse 7.b]. Le
passage de la cellule à l’organisme est bien une transformation qualitative
[thèse 9.b]. Une stratégie de lutte de l’organisme contre sa mort en tant
qu’organisation matérielle : La reproduction. Celle-ci accomplit une
conservation de la qualité de la matière vivante au niveau pluricellulaire, malgré
la mort individuelle. Nous connaissons de nombreux cas de reproduction
végétative ; la sporulation, le bouturage naturel, la scissiparité, … Mais
c’est la reproduction sexuée qui, en dernière instance, supplante toutes les
autres. Qu’elle soit sexuée ou asexuée, la reproduction correspond toujours à
une reconstruction ontogénétique à partir d’un cellule, ou d’un nombre
restreint de cellules.
Notons qu’à la différence de la reproduction cellulaire conforme, il
faut être deux pour accomplir la reproduction sexuée. Les deux qualités n’en
restent pas moins comparables dans le cadre de leur résultat (conservation
structurale dans le temps), en signalant que la reproduction sexuée est une
innovation imputable aux unicellulaires autant qu’aux pluricellulaires :
Les unicellulaires pratiquent dans leur immense majorité la reproduction
conforme et la reproduction sexuée au cours de leur cycle de
développement. Il faut envisager cette dernière comme une stratégie
supplémentaire de lutte pour l’existence dont le statut initial sera le même
pour les uni- et les pluricellulaires.
Mais l’organisme doit également lutter contre la désorganisation au
cours même de son existence. Désorganisation d’origine à la fois interne et
externe à l’organisme, qui a pour résultat ce que nous appellerons usure ou
vieillissement. Au final et conformément à ce qui vient d’être dit, la mort de
l’organisme peut aussi bien résulter d’une dégénérescence (morts cellulaires)
que d’une prolifération anarchique des tissus (cellules cancéreuses à extension
spontanée sur l’ensemble de l’organisme), puisqu’elle consiste en une
désorganisation triomphante : A ce niveau, la lutte consiste donc elle
même à limiter la mort des cellules mais aussi leur prolifération anarchique.
Cette dernière est comprise comme une désorganisation non-totale des propriétés
conservatrices au niveau cellulaire ; la spécialisation cellulaire
disparaît, la reproduction conforme subsiste : Le cancer est une
régression (désorganisation) des cellules du stade de conservation
pluricellulaire au stade moins évolué de conservation strictement cellulaire.
On situera la lutte contre l’usure sur trois niveaux de régulation (en
citant des exemples connus pour notre espèce) :
-
Propriétés régénératrices
(hémostase-cicatrisation, régénération d’organes ou de tissus, minéralisation
osseuse, renouvellement cellulaire),
-
Propriétés homéostatiques
(maintien de la température interne, de la glycémie, des équilibres hydriques,
salins et acido-basiques, sous l’effet d’interactions neuro-hormonales et
hormonales),
-
Propriétés immunitaires (dirigées
contre les agressions –vivantes ou non- de l’environnement mais aussi contre la
désorganisation interne –cancéreuse par exemple).
La conjonction de ces trois propriétés physiologiques assure le maintien
tendanciel de la structure pluricellulaire. Cependant, comme on s’en doute,
l’organisme trouvera dans ces processus dégénératifs / prolifératifs contre
lesquels il lutte des aspects concourrant à la conservation. En d’autres
termes, il retournera contre la désorganisation ses propres armes : Contre
la dégénérescence des tissus, la prolifération (mouvement de conservation) ;
contre le fléchissement de certaines conditions physico-chimiques internes, la
surproduction régulatrice (mouvement de compensation) ; contre
l’attaque d’un pathogène, la prolifération des cellules immunitaires et le
destruction des cellules infectées (mouvement de protection) ;
contre les déformations létales (au niveau des cellules tissulaires) liées à
l’interaction dynamique organisme – environnement, la prolifération
cellulaire –à valeur adaptative à l’échelle de l’organisme- telle que les
callosités cutanées, le « bronzage », le renforcement musculaire (mouvement
d’adaptation).
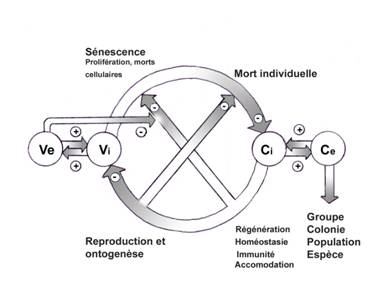
Voici donc, pour récapituler, cinq
processus spécifiques du niveau pluricellulaire (fig.3). Deux processus
désorganisateurs : Le vieillissement (par dégénérescence ou par
prolifération) et la mort. Deux processus conservateurs : L’activité
physiologique au sens large et la reproduction sexuée. Plus un processus
désorganisateur externe, indistinct dans le cadre d’une telle généralisation,
des processus désorganisateurs internes.
Première contradiction
Les mécanismes de stabilisation oeuvrent dans le sens d’une lutte contre
les morts cellulaires et les proliférations anarchiques en favorisant (en
contrôlant) des proliférations cellulaires mais aussi des morts cellulaires
locales. Deux exemples.
L’acquisition d’une aptitude à distinguer les molécules du soi
des molécules du non-soi (éventuellement pathogènes) passe par une destruction
des cellules immunitaires auto-réactives préalable, notamment au niveau du
thymus.
Les formes de l’organisme apparaissent au cours de l’embryogenèse :
Par exemple la main humaine embryonnaire est d’abord un « moufle »
avant que les doigts ne se dégagent du fait de la mort des cellules
intercalaires, les futurs oviductes et spermiductes coexistent avant de
dégénérer sélectivement, conformément au
sexe génétique.
Constatation simple : Ces mécanismes s’opposant à la
désorganisation en utilisant les mêmes armes qu’elle, contribuent de fait à faire
changer structuralement l’organisme pour qu’il soit mieux protégé des
fluctuations du milieu : Evolution de la morphologie en fonction de
l’activité physique, évolution du phénotype immunitaire, etc.
Deuxième contradiction
La reproduction est l’occasion pour l’organisme de perpétuer sa
structure à travers le temps et contre la mort individuelle. Là encore un avantage
collatéral se déclare…
La reproduction, sexuée ou non, est également une modalité spatiale dans
le sens où elle est « l’origine » des populations de même espèce
coexistant dans un même milieu. En tant que superstructure, l’ensemble des
relations sociales entre individus de l’espèce est une modalité externe à
chaque organisme, favorisant l’intégrité structurale de chaque organisme.
Citons le cas limite de
On distingue bien sûr plusieurs niveaux d’évolution de cette
superstructure, des espèces à individus quasiment isolés[9]
(modalité quasi-absente) aux espèces « sociales » (modalité évoluée
au plus haut degré chez les insectes sociaux[10]).
La superstructure proposée crée les conditions d’un changement
qualitatif du niveau de l’organisme au niveau plus complexe de la société
d’organismes. Sachant que le cannibalisme[11]
est dans la nature un phénomène particulièrement rare (même si les
néo-darwiniens surenchérissent souvent sur ces anecdotes si facilement
transposables à l’économie de marché !), indiquons ici que les écologistes
parlent indistinctement d’effet de groupe et de coopération pour les
populations uni- et pluricellulaires.
De plus, il est tout à fait finaliste –et anthropocentriste !- de
considérer la ruche ou la fourmilière comme la forme sociale en puissance de la
future Humanité ! Du point de vue marxiste, il serait hâtif de saisir
l’ensemble des espèces vivantes sans tenir compte de l’histoire particulière de
chacune. En l’occurrence, nous verrons que l’Humanité échappe partiellement aux
niveaux d’organisation supérieurs pour des raisons qui tiennent à son histoire,
notamment à ce que Patrick Tort appelle « l’effet réversif de
l’évolution » : Notre étude n’a pas du tout pour but de formaliser
une évolution nécessaire de toutes les formes de vie mais au contraire de
mettre à jour les processus évolutifs objectifs qui rendent possibles les
histoires nécessaires et nécessairement différentes de chaque forme de vie.
-
Contradiction antagoniste secondaire : Pour lutter contre la disparition imminente de sa structure,
l’organisme se reproduit. La reproduction sexuée est corrélativement la
condition de la formation d’une population spécifique partageant les mêmes
caractères et tirant bénéfices mutuels de leur superstructure : La
conservation produit donc une non-conservation : Transformation
qualitative organisme – espèce.
-
Contradiction antagoniste principale : Pour lutter contre sa désorganisation ante-mortem,
l’organisme dispose de propriétés régulatrices basées non sur l’inertie, mais
sur le terrain même de la désorganisation cellulaire. On peut d’ores et déjà
remarquer qu’à ce stade ;
·
La modalité de reproduction, fondée
sur des mécanismes cellulaires (donc d’ordre inférieur) impose, et c’est
nouveau, une ontogenèse produisant la structure par un certain type de
coopération cellulaire, avant d’avoir à la conserver (mouvement) : La
structure pluricellulaire , apparue dans le contexte d’une conservation
plus efficace des structures cellulaires, finit par nier cette tendance
initiale « au profit » de sa propre auto-conservation (prolifération
et/ou dégénérescence cellulaire).
·
La population, en tant que
superstructure du niveau de l’organisme individuel, favorise la conservation de
chaque organisme, mais peut finir par la nier pour assurer sa propre
auto-conservation (le « sacrifice » et plus généralement la tendance
à étendre par le peuplement le territoire occupé selon sa structure) :
Nous changeons alors de niveau…
Processus conservateurs et évolutifs liés à l’organisme
pluricellulaire : Ve ; modifications du milieu ou variations environnementales,
Vi ; modifications internes, Ci ; constance infra-structurale,
Ce ; Constance externe ou ultra-structurale.
Fig.3
C. Troisième niveau structural : Le groupe spécifique
L’enjeu fondamental de la biologie de l’évolution est depuis toujours
celui de mettre à jour les mécanismes de la spéciation, c’est à dire de la
création des espèces. Nous arrivons donc ici au cœur du problème.
Depuis quelques décennies, on a coutume de distinguer sans doute à
raison deux procès de spéciation: 1) la radiation à partir d’un type ancestral
spécifique d’un certain nombre d’écotypes, de variétés, de races, de
sous-espèces et consécutivement la transformation phylétique d’une
espèce ancestrale en un buisson d’espèces typologiquement apparentées (même
genre, même famille) correspondent à ce qu’on appelait la spéciation
darwinienne, en référence à ses travaux sur les différentes espèces de pinsons
des îles Galápagos, et qu’on appelle aujourd’hui micro évolution. 2)
Tout processus d’émergence historique d’un plan d’organisation totalement
nouveau à partir d’un plan ancestral comme les filiations poissons –
batraciens, reptiles – mammifères, etc. se rapporte en revanche au concept de macro
évolution.
Conceptuellement, la question de la définition de l’espèce a une
histoire complexe ; elle est loin d’être résolue aujourd’hui… La question
se résume à ceci : L’espèce a t-elle une réalité objective
(discontinuisme) ou n’est-elle qu’un concept trompeur imputable à notre vision
subjective du monde ? On pourrait penser le problème soluble par
l’expérimentation, ce n’est pas le cas. Position communément admise ; une
espèce est un ensemble d’individus interféconds (deux individus qui ne sont pas
interféconds appartiennent donc à deux espèces distinctes, même s’ils partagent
un phénotype quasi-similaire).
A l’évidence, si cette distinction passe nécessairement par la
reproduction sexuée, qu’en est-il donc de deux mâles de même espèce ?
Qu’en est-il des individus stériles ? Qu’en est-il des espèces qui ne pratiquent
que la reproduction asexuée ? Qu’en est-il enfin des espèces données comme
distinctes alors qu’elles sont encore interfécondes in vitro ?
La théorie synthétique, dans sa formulation récente, modère fortement
cette définition, mais pour lui préférer une définition pour le moins vague et
problématique, de l’aveu même de leurs auteurs. La raison de ce hiatus est
simple : Historiquement le discontinuisme est un argument fixiste, celui
de la typologie. Chaque espèce connue correspond à un type précisément défini,
il n’existe pas de populations intermédiaires, hybrides, en dégradé, entre deux
espèces connues, donc le transformisme n’est qu’une séduisante abstraction. On
remarquera que la théorie synthétique tend néanmoins à se départir de ses
positions traditionnellement continuistes (elles étaient celles de Darwin) au
profit d’un nouveau discontinuisme proposé par les saltationnistes cette fois[12].
Vue la position adoptée ici, celle d’une distinction qualitative qui
existe objectivement entre une cellule et une autre, entre un organisme et un
autre, posons que des distinctions qualitatives objectives existent entre les
espèces que l’Homme du reste distingue spontanément depuis toujours. Partant,
1) la thèse idéaliste voudra que chaque espèce ayant son type propre distinct
des autres, possède une essence immuable, trop parfaitement adaptée à
son milieu pour que le moindre changement
ne soit pas
mortel, 2) la thèse
matérialiste discontinuiste avancera
ceci : Une espèce
rassemble continûment, sur la
base de la reproduction sexuée, des
races inter-fécondes d’origine commune ; micro évolution unanimement reconnue, même
par les fixistes actuels, ces derniers ne critiquant
que la macro évolution. La distinction (barrière reproductive[13])
ne peut être qu’un événement (saut qualitatif) et non un fait surnaturel et
permanent, survenant dans le prolongement de la raciation. Cherchons donc
dialectiquement la clef d’un tel événement.
Au préalable il convient de remarquer, toujours à l’appui de notre
conception, qu’à l’instar de la cellule au sujet de laquelle nous assistions au
passage du caractère fortuit des modalités fondamentales de conservation à leur
caractère secondairement actif, téléologique, raffiné [voir annexe
« variabilité génétique et structures compensatrices »], nous
assistons ici encore, en apparence, au
passage du
caractère
contingent de la raciation au caractère nécessaire, activement
conservateur de la
spéciation : des mécanismes assureraient l’émergence des barrières interspécifiques[14] ;
Mais dans quel but ? La transformation d’un type spécifique en un
autre serait-elle une modalité de la conservation du type ?
Double contradiction
Conformément à la démonstration exposée pour les stades antérieurs
(c’est à dire pour les structures inférieures), considérons l’espèce comme une
entité vivante, c’est à dire comme une structure assiégée par l’inéluctable
mouvement de modification du type (ensemble des caractères
anatomo-physiologiques communs aux individus de l’espèce) et luttant contre lui
par un mouvement conservateur (perpétuation du type dans le temps).
Les données premières sont les suivantes : Une espèce naît, se
maintient pendant quelques dizaines de millions d’années en moyenne, puis
s’éteint. On s’intéressera d’abord à la manière dont l’espèce se maintient
pendant cette durée, se renouvelant de génération en génération par
reproduction sexuée, conformément à son type.
Chacun sait qu’une population artificielle de clones (ensemble
d’individus tous identiques génétiquement) s’avère extrêmement fragile face à
la moindre fluctuation de l’environnement : Pour ne citer qu’un exemple,
un champs de plantes issues d’un même clone sera instantanément exterminée par
la première infection virale.. Tout lycéen apprend en biologie qu’une espèce
déterminée par un ensemble de caractères spécifiques se compose
d’individus aux formes variées, autrement dit et pour employer le terme
consacré, que toute espèce est polymorphe. Pour chaque caractère on trouve au
sein de la population un certain nombre de formes (par exemple pour le
caractère pigmentation des yeux, chez l’Homme, on connaît plusieurs
formes ; les couleurs).
Le polymorphisme d’une espèce ne remet pas en cause son type, il
consiste au contraire à produire le plus de versions possibles du type sans le
remettre en cause, c’est à dire à produire une variabilité maximale de
potentialités fonctionnelles (de survie en conditions changeantes) à partir
d’un type donné. Par exemple, la diversité des teintes de coquilles chez une
espèce d’escargots permet a priori une adaptation potentielle plus grande (camouflage) à
des environnements différents (supports colorés comme le
sable, l’humus, etc.). Très clairement, l’avantage du
polymorphisme est de
favoriser telle ou telle forme représentée a priori lorsque le
milieu change, donc d’assurer autant que faire se peut la perpétuation de
l’espèce malgré les fluctuations du milieu : Il constitue une lutte
« conservatrice » contre les macro mutations
« monstrueuses » viables.
Précisons au passage que le concept de type est entendu ici non comme
essence cachée de l’organisation anatomo-physiologique mais comme cette
organisation elle même, contenant en germe sa propre négation.
On pensera d’abord que cette modalité conservatrice au niveau de
l’espèce résulte des mutations génétiques, c’est à dire d’évènements fortuits.
Cette hypothèse est en partie vérifiée : Toutes les versions
(allèles) d’un
gène résultent effectivement d’une
accumulation historique de mutations rares. Cependant le polymorphisme apparaît comme un processus
actif, ayant pour base les mutations, intervenant au niveau de
la reproduction sexuée[15]
et réitérée à
chaque génération, contre
une homogénéisation éventuelle
en milieu « non-changeant » (à notre échelle de perception du temps).
Tous les néodarwinistes s’accordent à dire que le polymorphisme est la
base minimale de l’évolution des espèces, au sens où il permet la formation de
groupes distincts par leurs formes, variétés, races puis espèces distinctes.
Puisque les variations sont endogènes et autonomes (mutations), comment
expliquer que les espèces soient dans leur structure typologique si bien
adaptées à leur environnement ? C’est là en général qu’intervient la
sélection naturelle. Les néodarwinistes lui donnent la définition
suivante : L’individu porteur d’une mutation « favorable »
(avantage sélectif) sera plus fécond que ses congénères ; ainsi ce gène
muté, ce nouveau gène, ne finira par dominer dans la population qu’après de
nombreuses générations (reproduction sexuée). Au passage, nous sommes
aujourd’hui bien loin de la notion tautologique de « survie du plus
apte »[16] dont le « bons
sens » a commis les ravages idéologiques que l’on sait. Cependant le
concept, qu’il faut semble t-il conserver coûte que coûte, reste bien
mystérieux, aux yeux même des néodarwiniens : Comment un changement de
couleur des yeux peut-il conférer à leur propriétaire une fécondité plus grande
(augmentation du nombre de descendants) ? A l’évidence la sélection
naturelle est le talon d’Achille du néodarwinisme, tous leurs dissidents ne se
sont pas privés de le dire. Mais c’est précisément en tant que « moteur de
l’évolution » que la sélection naturelle porte préjudice à la solidité du
néodarwinisme !
Laissons de côté ce concept si agréable aux matérialistes mécanistes
[voir annexe « la question de l’hérédité des caractères acquis »] et
revenons au problème du polymorphisme. Forcé
de la considérer comme le
résultat d’un phénomène actif plutôt que comme l’état passif d’une population
assiégée par des mutations accidentelles permanentes, tout savant sait que son
origine est la méiose.
Une fois encore, la méiose est un processus nécessaire dont la vocation
première n’est pas à première vue le polymorphisme de la population mais la
fécondation formant une cellule-œuf, raison d’être de la reproduction sexuée.
En marxiste, remettons les choses à l’endroit. Considérée au niveau de
l’unicellulaire ou de l’individu pluricellulaire, la reproduction sexuée n’a
absolument aucun avantage conservateur sur une reproduction asexuée : Les
deux types de reproduction résolvent semblablement le problème de la
perpétuation dans le temps… Ce qui est « neuf » dans l’œuf, c’est la
combinaison particulière des formes, non les formes elles-même, qui existent
déjà dans la population isolément les unes des autres : Diversifier les combinaisons sans
changer les formes,
c’est, nous l’avons
dit, protéger le
type général des
fluctuations du
milieu dans lequel il est adapté[17].
Alors qu’on nous apprend que la méiose est une condition permettant aux
espèces diploïdes[18]
de pratiquer la reproduction sexuée, avançons au contraire que la diploïdie est
une condition permettant aux espèces de pratiquer une reproduction
sexuée (dont la méiose fait
partie intégrante), c’est
à dire de pratiquer une diversification favorisant
sa conservation en tant que type. Les généticiens des populations nomment cette
propriété « avantage du poly-allélisme »…
On peut souligner ainsi le raffinement du processus méiotique : Il
s’agit de produire à partir d’une combinaison donnée de versions géniques
(génotype d’un individu, identique pour toutes ses cellules) une
« infinité » de cellules sexuelles génétiquement différentes (nombre
maximal de combinaisons différentes de ces versions géniques appelées
allèles) : C’est un tri qui retiendra pour chaque gène du génotype
parental une version sur deux (l’ensemble de ces versions se retrouvant
ensemble dans une cellule sexuelle correspondra à une combinaison quasi unique statistiquement).
Chaque cellule sexuelle est haploïde (une seule version de chaque gène, puisque
chaque chromosome n’y est plus présent qu’en un unique exemplaire (maternel ou
paternel), de sorte que lors de la fécondation, la réunion dans la cellule-œuf
des deux noyaux rétablira la diploïdie de l’espèce : Le cycle de
développement de l’espèce est bouclé.
Problème : Les gènes ne sont pas matériellement libres les uns des
autres dans le noyau d’une cellule. Ils sont liés les uns aux autres en un
certain nombre de groupes de gènes, les chromosomes. Chaque chromosome existant
en double exemplaire dans la cellule diploïde (2 fois 23 chromosomes = 46
chromosomes chez l’être humain par exemple) : La disjonction des
chromosomes homologues au moment de la méiose (formation de cellules haploïdes
à partir d’une cellule diploïde) permet déjà de produire potentiellement un
grand nombre de combinaisons chromosomiques haploïdes différentes (brassage
inter-chromosomique) : Plus le caryotype d’une espèce contiendra de chromosomes,
plus la diversité génétique des gamètes sera grande.
A ce brassage simple et déjà avantageux pour la variabilité des gamètes
s’ajoute un brassage beaucoup plus raffiné, connu sous le nom de brassage
intra-chromosomique. Il s’agit de lever le problème résiduel de la liaison
physique des gènes qui sont situés sur un même chromosome. Avant de se
disjoindre, deux chromosomes homologues s’enjambent en plusieurs points
aléatoirement, puis s’échangent des segments entiers d’ADN ; Des versions
génétiques différentes situées initialement sur des chromosomes homologues
distincts peuvent donc se retrouver en fin de méiose sur le même chromosome recombiné.
La diversité des combinaisons géniques s’en trouve considérablement augmentée
au niveau des cellules sexuelles.
Nous n’entrerons pas dans les détails moléculaires très complexes d’un
tel processus de brassage intra-chromosomique. Etonnons nous simplement, une
fois de plus, du raffinement extrême de cette « stratégie » de
diversification !
Le discontinuisme impose qu’une barrière reproductive (mécanismes
cellulaires perfectionnés empêchant par exemple la fécondation d’un ovule par
un spermatozoïde d’une autre espèce) s’érige à un moment donné entre un groupe
d’individus et sa population d’origine (spéciation). Ce moment n’a jamais été
observé, du moins pour les espèces actuelles, et les mécanismes éventuels du
processus même restent énigmatiques[19].
Citons seulement des cas de processus actifs d’incompatibilité spécifique entre
de nombreuses plantes à fleur et du pollen d’espèce distincte : Le rejet
du pollen incompatible est ici effectué non par l’ovule à féconder mais par le
pistil (organe reproducteur contenant les ovules). En ce sens, on voit à quel
point les enjeux de cette barrière reproductive se situent moins à l’échelle
cellulaire qu’à l’échelle pluricellulaire.
Les mécanismes, sans doute tout à fait perfectionnés, d’isolement
reproductif, s’inscrivent donc en creux dans notre analyse, en particulier pour
le règne animal (les incompatibilités semblent chez eux beaucoup plus
complexes, mais se situent sans doute autant sinon plus au niveau
inter-individuel – phéromones, désynchronisation des périodes de reproduction
entre espèces, incompatibilités anatomiques -
qu’au niveau intercellulaire – non-reconnaissance ovule/spermatozoïde,
non développement de l’œuf en cas de fécondation accidentelle - ). Les enjeux
ne font quant à eux aucun doute : La raciation est la négation de la
conservation typologique, positivement préparée par cette dernière
(polymorphisme), la spéciation est la négation de cette négation (une barrière
reproductive réinstalle le processus conservateur sur un type nouveau).
Effet collatéral évident de ce mécanisme : La multiplication des
espèces crée les conditions de formation de structures d’ordre supérieur, les
écosystèmes. Que l’écosystème, structuré par des symbioses, des relations
trophiques, des coadaptations, régulant quantitativement et qualitativement les
populations qui le composent, soit à l’évidence et de l’avis même des
écologistes une structure conservatrice (même si elle évolue souvent par stades
sur plusieurs siècles : exemple typique des tourbières), légitime à son
niveau cette négation de la négation qu’est la spéciation au niveau inférieur
(distinction d’espèces qualitativement différentes pour que des relations
trophiques puissent avoir lieu entre elles, en vue de maintenir les
écosystèmes).
-
Contradiction antagoniste secondaire : La reproduction sexuée et, à travers elle la méiose, permettent
aussi une lutte contre l’extinction de l’espèce par inadaptation collective aux
conditions changeantes du milieu, l’espèce conserve son type en le rendant le
plus polymorphe possible. Ce polymorphisme est pourtant la base d’une
micro-évolution éloignant progressivement les populations successives du type
moyen. Pourquoi cet éloignement ? Parce que le type n’est justement pas
une essence ! Loin de nous donc l’idéalisme typologique du XIXe
siècle.
-
Contradiction antagoniste principale
(fig.4) : La reproduction sexuée contribue à
travers toute une série de processus raffinés, de la méiose conservant le
caryotype à la barrière reproductive en passant par les mécanismes
d’histo-compatibilité limitant les risques d’autogamie, à entraver l’apparition
de formes trop différenciées les une des autres. Pourtant ces
différenciations peuvent aboutir à des
distinction phénotypiques avantageuses pour la survie de l’espèce (dimorphisme
sexuel chez la plupart des animaux, castes des insectes sociaux, division du
travail, etc.), autant qu’à des macro mutations profondes et négatrices ;
les phénomènes de néoténie, d’apomorphose, etc., liés aux remaniements
chromosomiques et impliquant des bouleversements ontogénétiques, assez
fréquents dans la nature (voir Les horloges du vivant, Jean Chaline).
Processus conservateurs et évolutifs liés au groupe spécifique : Ve ; modifications du milieu ou variations environnementales,
Vi ; modifications internes, Ci ; constance infra-structurale,
Ce ; Constance externe ou ultra-structurale.
Fig.4

D. Quatrième niveau structural : La biosphère
C’est une discipline assez jeune, l’écologie, qui a élaboré la notion de
biosphère. Notion dont les vertus explicatives sont en apparence restreintes au
champ de l’écologie ; Il n’est jamais question de biosphère en matière
d’évolution darwinienne, mais il est question d’évolution (au sens large) en
matière d’environnement.
On cherche à montrer en particulier, pour des raisons de prévention
écologique, que l’environnement est aujourd’hui en équilibre fragile, que cet
équilibre résulte d’une interaction entre histoire de
Définition courante : La biosphère correspond à la totalité de la
matière vivante (aquatique et aérienne, animale et végétale, uni et
pluricellulaire) couvrant la surface terrestre, sorte de pellicule de vie plus
ou moins continue, en interaction avec l’hydrosphère, l’atmosphère (échanges
gazeux) et la lithosphère (actions directes ou indirectes sur l ‘érosion,
la formation des calcaires et des combustibles fossiles, …).
Paradoxalement, ce sont les géologues et les climatologues qui ont forgé
ce concept, en dehors du champ de la science de l’évolution. Il est pourtant
clair que cette biosphère a une histoire ; L’histoire de
► Formation
de matière organique océanique (molécules « pré-biotiques »), encore
appelée « soupe primitive ». L’atmosphère est initialement très riche
en dioxyde de carbone (gaz à effet de serre) et totalement dépourvue de
dioxygène.
►
Apparition des premières cellules vivantes, qui consomment ces molécules pour
1) renouveler leur propre matière organique (= matière première), 2) produire
de l’énergie utilisable par fermentation (= carburant). La vie aérienne est
impossible (trop d’ultraviolets, trop de dioxyde de carbone favorisant un
puissant effet de serre).
► Les
molécules organiques de la soupe primitive viennent à manquer, mais la
biosphère, exclusivement unicellulaire, aquatique et extrêmement ténue,
subsiste grâce à une innovation de
grande
importance :
La photosynthèse. De nombreuses espèces finissent par se passer de la matière
organique de l’environnement (hétérotrophie) et deviennent autotrophes :
elles consomment du dioxyde de carbone et de l’eau (comme matière première), de
la lumière (comme source d’énergie) et rejettent massivement un déchet
métabolique (inutile pour elles) ; le dioxygène. Les océans puis
l’atmosphère s’appauvrissent en dioxyde de carbone (l’effet de serre baisse, le
climat s’en trouve modéré) et s’enrichissent en dioxygène (dont une partie
forme dans la haute atmosphère une couche d’ozone, ou trioxygène, absorbant une
grande partie des ultraviolets).
►
L’enrichissement atmosphérique en dioxygène plafonne lorsque les cellules
s’arment d’une nouvelle innovation de taille, beaucoup plus rentable que la
fermentation en matière de production d’énergie, la respiration cellulaire
(consommation de dioxygène et rejet de dioxyde de carbone) : Les
concentrations atmosphérique et hydrosphérique en dioxygène se stabilisent du
fait d’une « régulation » entre les intensités globales de la
photosynthèse et de la respiration biosphérique.
► La vie,
toujours aquatique, innove en se compliquant. Les espèces coloniales puis
pluricellulaires à différenciation apparaissent, dans le règne animal comme
dans le règne végétal.
► La
préexistence fortuite d’une couche d’ozone et l’extrême pauvreté de la teneur
atmosphérique en dioxyde de carbone rend la vie aérienne possible. Seule
difficulté ; le déficit hydrique des milieux aériens. Une partie de la
biosphère sort de l’eau : Chez les végétaux, on passe des algues aux
bryophytes (mousses, aériennes mais inféodées aux milieux humides) puis aux
spermatophytes qui dominent aujourd’hui sur Terre. Les arthropodes végétariens
semblent être les premiers invertébrés à conquérir la terre ferme. Chez les
animaux vertébrés, on passe des poissons aux batraciens (aériens inféodés aux
milieux humides) puis aux reptiles, oiseaux, mammifères (parmi les très
nombreuses innovations délivrant ces animaux des problèmes liés au déficit
hydrique, citons celles de l’homéothermie, de la ventilation pulmonaire, de
l’œuf à coquille puis de la viviparité).
L’image d’Epinal marquant les esprits dans ce scénario est celle du
premier « poisson à pattes » sortant un jour de l’eau pour habiter un
désert aride… Quoi de plus absurde ? La conquête du milieu aérien est
inconcevable à notre échelle (d’individus), pourtant elle a eu lieu. Ce ne sont
ni quelques individus, ni même quelques espèces qui sont « sortis »
de l’eau, mais des écosystèmes, seules entités capables d’une certaine
autonomie vis à vis du biotope (conditions physiques du milieu) [autonomie et
indépendance n’ont pas la même signification !]. Il y a fort à parier que
ces derniers aient d’ailleurs été précédés non d’écosystèmes océaniques mais
d‘écosystèmes continentaux d’eau douce (étape intermédiaire, requerrant des adaptations
propres).
Pourquoi insister sur cette notion d’écosystème ? Parce que nous
changeons ici de niveau structural. Une seule espèce et encore moins un seul
individu ne peut déterminer les sauts qualitatifs de l’histoire biosphérique.
Voilà pourquoi la notion de biosphère revêt ici plus qu’une vertu descriptive,
cantonnée à la climatologie ; Sa valeur concrète, objective, achève notre
exposé par un stade, ultime certes, mais toujours en mouvement, celui de la
macro-évolution.
Première contradiction
Ce scénario tracé à grands traits peut être développé en chacune de ses
étapes chronologiques grâce aux nombreux documents fossiles recueillis depuis
plus d’un siècle : Il consiste, pour telle partie du monde où les fossiles
se sont empilés chronologiquement dans les sédiments, en une succession
discontinue de faunes et de flores différentes. On a coutume de citer, pour
prendre l’exemple le plus connu, la succession Ere des reptiles et des
conifères (Secondaire) – Ere des mammifères et des plantes à fleurs (Tertiaire).
Succession discontinue parce que ponctuée par une catastrophe planétaire
(météorite et/ou volcanisme intense) dont l’existence est communément admise
–événement on ne peut plus contingent-. Cependant, on admet aussi communément
aujourd’hui, que la diversité des reptiles du secondaire était en déclin bien
avant l’instant de cette catastrophe.
De tels remplacements faunistiques et floristiques peuvent sembler très
schématiques ou subjectifs (état dégradé des documents fossiles). Gardons-en
néanmoins le cadre conceptuel, lui aussi communément admis ; Ces
remplacements ponctués par ce que les paléontologues ont souvent appelé sans
doute à tort des catastrophes, dont les modalités sont du reste rarement
établies, sont nombreuses au cours de l’Histoire de
On connaît de
plus dans l’histoire de la biosphère un événement qui reste inexpliqué, que ce
soit dans le cadre catastrophiste ou dans un autre (la théorie des extinctions
de J.M.Raup par exemple, chapitre 3) : La « crise » cambrienne
n’est marquée par aucune extinction massive mais par une diversification d’une
puissance inégalée par la suite[20].
La succession des crises apparaît donc peu contingente.
A l’instar d’une espèce qui en produit d’autres, apparentées, avant de
s’éteindre (conservation négatrice dans le temps), un ensemble d’espèces (en
inter-relations) dominé par tel ou tel plan d’organisation anatomique produit
de nouveaux ensembles, minoritaires et marqués par de grandes innovations avant
de s’éteindre massivement. On préférera donc expliquer les crises par un mouvement
dialectique (qualitatif et endogène) plutôt que par un mouvement mécanique
(brutal, contingent et exogène) en remarquant 1) l’antériorité de l’émergence
macro-évolutive sur l’extinction massive, 2) la fréquence des crises au cours
de l’Histoire de
L’étonnante communauté typologique d’une classe d’être vivants (par
exemple : Tous les oiseaux possèdent un plumage, des ailes, des sacs
aériens, un bec corné, …) impose une distinction qualitative entre
micro-évolution (spéciation) et macro-évolution (émergence d’un plan
d’organisation radicalement différent). Cette distinction, irritante pour les
gradualistes, était un des principaux contre-arguments fixistes (voir
Denton) ; Elle est à présent un argument évolutionniste dialectique.
Deuxième contradiction
Au cours de la période de domination d’une classe d’êtres vivants, c’est
à dire avant la crise, conçue comme une désapdaptation inéluctable du plan
d’organisation des classes coexistantes dans certains milieux (désadaptation
ayant pour origine l’évolution de l’environnement et/ou le
déficit d’évolu-
tion de ces
classes), celle-ci conserve son plan général d’organisation en favorisant la
multiplication des versions originales de ce plan. Cette multiplication
s’intègre à l’évidence dans le processus macro-évolutif : L’autruche et le
pinson sont radicalement différents, bien qu’appartenant tous deux à la classe
des oiseaux.
La macro évolution infra-classe diversifie un type de façon à multiplier
les adaptations générales au milieu : de nombreux poissons comme l’anabas,
le périophtalme, les dipneustes, sont ainsi adaptés à la vie
aérienne en conservant leur appartenance à la classe des poissons. La conquête
de nouveaux milieux par macro mutation infra-classe est un mode de conservation
de la classe contre les excédents ou les manques relatifs à un milieu donné,
saturé par la micro évolution. En se prolongeant dans le milieu conquis, la
macro évolution dépasse sa vocation à conserver un type général et devient
« super-classe », orientant le nouveau plan d’organisation dans le
sens d’une meilleure adaptation au milieu : Les poissons forment ainsi le
groupe ancestral des batraciens, les batraciens celui des reptiles, les
reptiles celui des oiseaux et des mammifères, toujours dans le sens d’un
indépendance de plus en plus marquée vis à vis du milieu aquatique (viviparité,
homéothermie, ventilation pulmonaire, excrétion, …).
Voici donc deux contradictions étroitement associées (fig.5),
visiblement fondées sur le même principe conservation/diversification. La
macro-évolution super-classe est conçue à la fois comme l’échec de la
macro-évolution infra-classe (conservation du type général de la classe en
diversifiant ses familles, groupes, ordres) et comme la réussite de la
conservation à un plus haut niveau de la représentation des êtres vivants sur
Terre (biosphère).
Avantage
collatéral : Cette macroévolution qui réadapte le plan d’organisation
des espèces en milieu changeant (conservation de la vie dans le temps) peut
fortuitement adapter ce plan à un autre milieu contemporain du premier
(extension spatiale de la biosphère) grâce à des innovations
anatomo-physiologiques (complexification). C’est dans ce sens qu’on peut
comprendre la colonisation de nouveaux milieux par la biosphère durant son
histoire : On peut considérer qu’aujourd’hui la majorité des habitats,
même les plus « hostiles » (fortes pressions, fortes températures,
faibles températures, forte acidité, obscurité totale, aridité extrême,
anaérobiose) est peuplée.
Perpétuée dans le temps (trois milliards d’années) et dans l’espace (la
quasi-totalité de la surface terrestre), la biosphère est confrontée au
changement global de la planète. L’avantage récemment mis à jour tient au fait
que les diverses interactions régulatrices (stabilité des concentrations
atmosphériques en dioxygène et en dioxyde de carbone par exemple) actuelles
tendent à stabiliser l’environnement planétaire (toujours sans le pouvoir
absolument !). Une certaine indépendance vis à vis du milieu se révèle (au
niveau de la biosphère, pas à celui des structures inférieures), tendant à
harmoniser (sans le pouvoir) les mouvements internes et externes à la matière
vivante. L’écologiste constatera aujourd’hui, ce n’était pas le cas au début de
l’histoire de
La diversité des espèces, se régulant quantitativement et
qualitativement au sein d’un écosystème, tend à pérenniser chacune d’elles,
sans le pouvoir à long terme (contradiction antagoniste) : On peut encore
considérer l’écosystème (dont l’extension est la biosphère) comme une
superstructure favorisant la conservation du niveau inférieur des groupes
spécifiques mais aussi l’émergence d’une structure hautement complexe et douée
de puissantes facultés conservatrices ; la biosphère.
-
Contradiction antagoniste
secondaire : La macro-évolution super-classe est le processus grâce auquel
la biosphère traverse ses « crises » biologiques en changeant ses
formes dominantes. Elle rend également irréversible l’installation de la vie
sur les milieux conquis par la macro-évolution infra-classe.
-
Contradiction antagoniste
principale : La macro-évolution infra-classe équivaut à un
« polymorphisme d’espèce » : Elle diversifie les familles et les
ordres pour conserver le type de la classe (conjonction de caractères corrélés
les uns aux autres[21]).
Une telle diversification produit des versions efficaces mais aussi des échecs
nécessaires (au sens où une forme plus complexe est un échec pour la
conservation temporelle de la forme simple). Ces derniers, plans d’organisation
radicalement nouveaux, caractérisant par exemple l’apparition d’une nouvelle
classe ou d’un nouvel embranchement, sont susceptibles d’être à leur tour
auto-conservateurs dans de nouveaux milieux terrestres.
Processus conservateurs et évolutifs liés à la biosphère : Ve ;
modifications du milieu ou variations environnementales, Vi ;
modifications internes, Ci ; constance infra-structurale, Ce ;
Constance externe ou ultra-structurale.
Fig.5
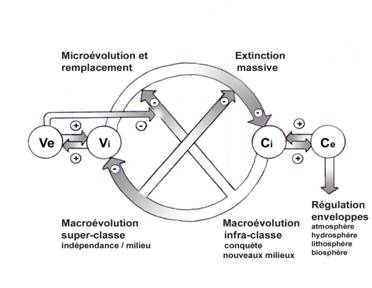
E. Conclusion
On le comprendra facilement, une telle synthèse ne peut être que
schématique. C’est volontaire. Son but avoué est de montrer en quoi le
matérialisme dialectique peut se loger légitimement et intégralement, sans
distorsion, au sein des éléments actuels de la théorie de l’évolution, de
montrer qu’il permet de penser les différents concepts connus des biologistes
dans leurs rapports objectifs. Tout reste à découvrir à partir de ce modèle,
lui même modifiable. Mais la description d’un phénomène n’est pas la
connaissance de ce phénomène. C’est une mauvaise habitude de biologiste
agnostique à laquelle il s’agit de tordre le cou…
On pourrait croire que la propriété conservatrice de la matière vivante
ressemble fort à un postulat vitaliste. Mais la vie n’incarne pas une force
surnaturelle « conservatrice » qui lui permettrait de survivre à
toutes les aventures terrestres en évoluant. C’est son histoire elle-même
qu’elle incarne. Et la propriété conservatrice qu’elle manifeste n’est pas la
cause de cette histoire, c’en est à la fois la cause et la conséquence.
[Cause et effet]
sont des concepts n’ayant comme tels une validité que s’ils sont appliqués à un
cas particulier, mais dans la mesure où nous considérons ce fait singulier dans
sa connexion générale avec la totalité du monde, (…) ils se confondent dans
l’action universelle réciproque, où cause et effet permutent continuellement
leur position ; Ce qui ici était effet devient ensuite cause et
vice-versa.
Engels. Dialectique de
C’est à la lumière de cette citation qu’il faut comprendre le problème
sous jacent à notre synthèse, celui de la conservation de la propriété
conservatrice elle-même… La théorie de l’hérédité qu’il est nécessaire d’établir
maintenant s’en trouve profondément compliquée, mais elle n’est pas
inaccessible…
Indiquons, et c’est un résultat global de l’exposé, que c’est
l’inefficience de cette conservation à un niveau donné qui détermine la
possibilité d’une conservation du niveau qui lui succède, sur les bases
procédurales non-perdues de conservation du niveau qui l’a précédé :
Paradoxalement donc, la perte non totale des mécanismes conservateurs à un
niveau (ceux des niveaux inférieurs persistent) –régression du mouvement conservateur-
établit positivement l’émergence des mécanismes conservateurs suivant le saut qualitatif à un niveau supérieur
–complexification du mouvement évolutif-.
► L’échec
de conservation architecturale de l’organisme pluricellulaire à différenciation
(B) se traduit au niveau de ses gamètes par une non-conservation du génome
(méiose), créant ainsi les bases du polymorphisme, dont le caractère
conservateur a été mis en évidence au niveau supérieur C de l’espèce sur les
bases des mutations génétiques et de la reproduction sexuée existant au niveau
inférieur A de la cellule.
► L’échec
de conservation typologique de l’espèce (C) se traduit par une micro-évolution
(édification d’une barrière reproductive dont les mécanismes, encore peu
connus, sont indéniablement relatifs à la physiologie de l’organisme –niveau
inférieur B-, créant ainsi les bases d’une conservation d’un plan
d’organisation sans type (macro-évolution infra-classe) pour lequel la loi de
corrélation des organes reste valable au niveau supérieur D (ensemble des
espèces « adaptées » les unes aux autres et à leur biotope ;
écosystème).
► L’échec
de conservation du plan d’organisation propre à une classe (D) se traduit par
une macro-évolution super-classe, condition du dépassement des crises
biologiques (conservation de la matière vivante au sens large) au niveau de la
biosphère, sur la base du polymorphisme (niveau inférieur C de l’espèce).
Pour souligner la réalité de l’action de ce niveau structural inférieur
C dans ce troisième point (espèce polymorphe, et non cellules) : Les gènes
« architectes » sensés définir au moment de l’embryogenèse
l’architecture générale de l’organisme (plan d’organisation), sont hautement
conservés (quasiment aucune mutation au cours du temps) à travers la plupart
des espèces animales : Par exemple les gènes architectes contrôlant la
construction des yeux sont à quelques rares mutations neutres près, les mêmes
chez la mouche et chez l’Homme (alors que leurs yeux sont absolument différents
dans leur structure !). En revanche la position et le nombre de ces gènes
diffère d’un phylum à l’autre.
Les recombinaisons de gènes semblent donc avoir un rôle déterminant sur
leurs effets, au moins aussi important que celui des mutations intra-géniques
dont on parle si souvent, qui sont d’ailleurs divers : Le même gène
architecte contrôle chez l’Homme à la fois la construction de la main à cinq
doigts et l’appareil uro-génital ! C’est un résultat peu orthodoxe aux
yeux des néodarwiniens mais néanmoins unanimement accepté aujourd’hui. Or les
processus relatifs au polymorphisme sont responsables de telles recombinaisons.